

The interaction between cold and light controls the expression of the cold-regulated barley gene cor14b and the accumulation of the corresponding protein
- PMID: 9952464
- PMCID: PMC32145
- DOI: 10.1104/pp.119.2.671
The interaction between cold and light controls the expression of the cold-regulated barley gene cor14b and the accumulation of the corresponding protein
Abstract
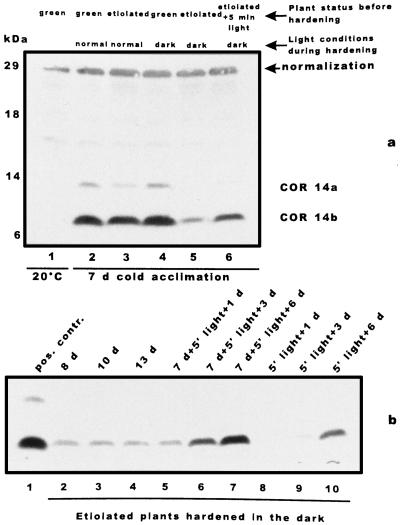
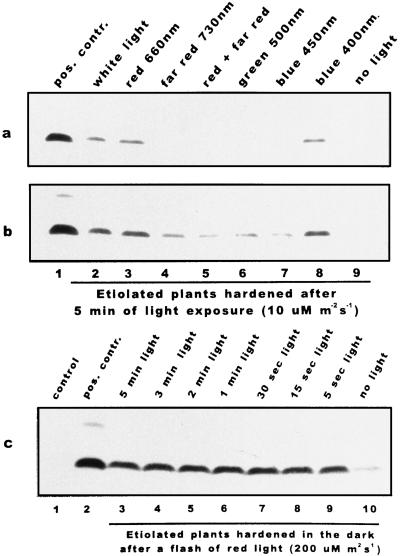
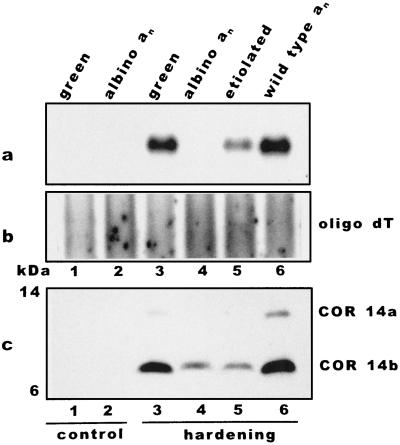
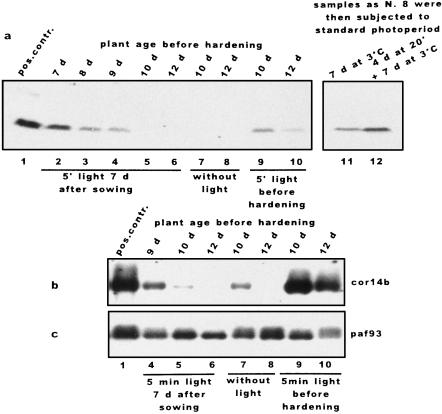
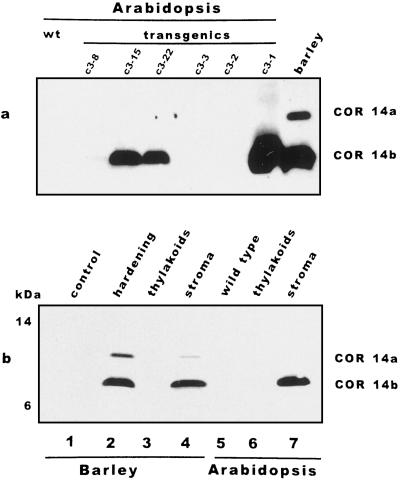
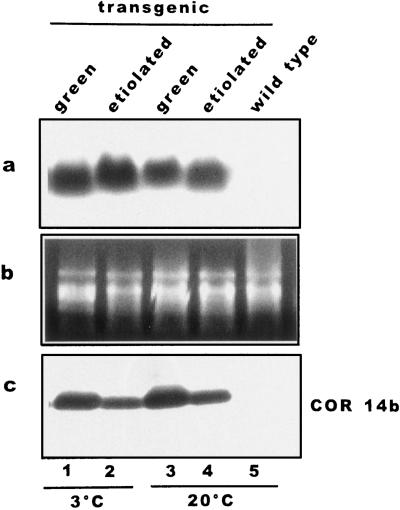
We report the expression of the barley (Hordeum vulgare L.) COR (cold-regulated) gene cor14b (formerly pt59) and the accumulation of its chloroplast-localized protein product. A polyclonal antibody raised against the cor14b-encoded protein detected two chloroplast COR proteins: COR14a and COR14b. N-terminal sequencing of COR14a and expression of cor14b in Arabidopsis plants showed that COR14a is not encoded by the cor14b sequence, but it shared homology with the wheat (Triticum aestivum L.) WCS19 COR protein. The expression of cor14b was strongly impaired in the barley albino mutant an, suggesting the involvement of a plastidial factor in the control of gene expression. Low-level accumulation of COR14b was induced by cold treatment in etiolated plants, although cor14b expression and protein accumulation were enhanced after a short light pulse. Light quality was a determining factor in regulating gene expression: red or blue but not far-red or green light pulses were able to promote COR14b accumulation in etiolated plants, suggesting that phytochrome and blue light photoreceptors may be involved in the control of cor14b gene expression. Maximum accumulation of COR14b was reached only when plants were grown and/or hardened under the standard photoperiod. The effect of light on the COR14b stability was demonstrated by using transgenic Arabidopsis. These plants constitutively expressed cor14b mRNAs regardless of temperature and light conditions; nevertheless, green plants accumulated about twice as much COR14b protein as etiolated plants.
Figures

References
-
- Adam Z. Protein stability and degradation in chloroplasts. Plant Mol Biol. 1996;32:773–783. - PubMed
-
- Bassi R, Dal Belin Peruffo A, Barbato R, Ghisi R. Differences in chlorophyll-protein complexes and composition of polypeptides between thylakoids from bundle sheath and mesophyll cells in maize. Eur J Biochem. 1985;146:589–595. - PubMed
-
- Bergantino E, Dainese P, Cerovic Z, Sechi S, Bassi R. A post-transcriptional modification of the photosystem II subunit CP29 protect maize from cold stress. J Biol Chem. 1995;265:8474–8481. - PubMed
-
- Bergantino E, Sandonà D, Cugini D, Bassi R. The photosystem II subunit CP29 can be phosphorylated in both C3 and C4 plants as suggested by sequence analysis. Plant Mol Biol. 1998;36:11–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
