

Infection process of the hepatitis B virus depends on the presence of a defined sequence in the pre-S1 domain
- PMID: 9971786
- PMCID: PMC104448
- DOI: 10.1128/JVI.73.3.2052-2057.1999
Infection process of the hepatitis B virus depends on the presence of a defined sequence in the pre-S1 domain
Abstract
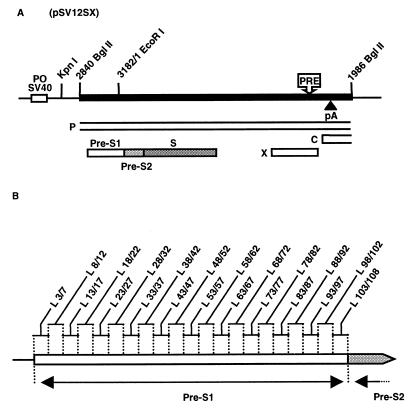
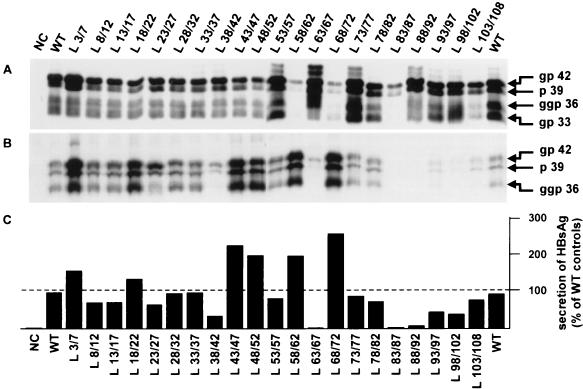
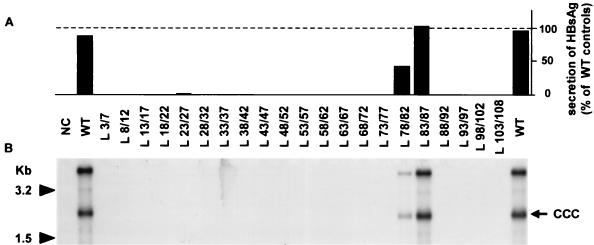
During the life cycle of hepatitis B virus (HBV), the large envelope protein (L) plays a pivotal role. Indeed, this polypeptide is essential for viral assembly and probably for the infection process. By performing mutagenesis experiments, we have previously excluded a putative involvement of the pre-S2 domain of the L protein in viral infectivity. In the present study, we have evaluated the role of the pre-S1 region in HBV infection. For this purpose, 21 mutants of the L protein were created. The entire pre-S1 domain was covered by contiguous deletions of 5 amino acids. First, after transfection into HepG2 cells, the efficient expression of both glycosylated and unglycosylated L mutant proteins was verified. The secretion rate of envelope proteins was modified positively or negatively by deletions, indicating that the pre-S1 domain contains several regulating sequences able to influence the surface protein secretion. The ability of mutant proteins to support the production of virions was then studied. Only the four C-terminal deletions, covering the 17 amino acids suspected to interact with the cytoplasmic nucleocapsids, inhibited virion release. Finally, the presence of the modified pre-S1 domain at the external side of all secreted virions was confirmed, and their infectivity was assayed on normal human hepatocytes in primary culture. Only a short sequence including amino acids 78 to 87 tolerates internal deletions without affecting viral infectivity. These results confirm the involvement of the L protein in the infection step and demonstrate that the sequence between amino acids 3 and 77 is involved in this process.
Figures

References
-
- Aden D P, Fogel A, Plotkin S, Damjanov I, Knowles B B. Controlled synthesis of HBsAg in a differentiated human liver carcinoma-derived cell line. Nature (London) 1979;282:615–616. - PubMed
-
- Bruss V, Hagelsten J, Gerhardt E, Galle P R. Myristylation of the large surface protein is required for hepatitis B virus in vitro infectivity. Virology. 1996;218:396–399. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
