

Functional domains of Tat required for efficient human immunodeficiency virus type 1 reverse transcription
- PMID: 9971835
- PMCID: PMC104497
- DOI: 10.1128/JVI.73.3.2499-2508.1999
Functional domains of Tat required for efficient human immunodeficiency virus type 1 reverse transcription
Abstract
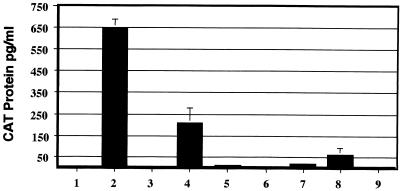
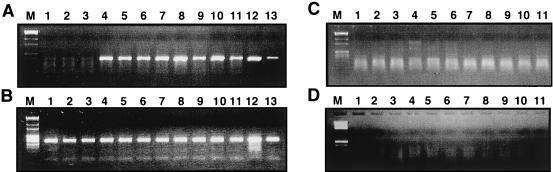
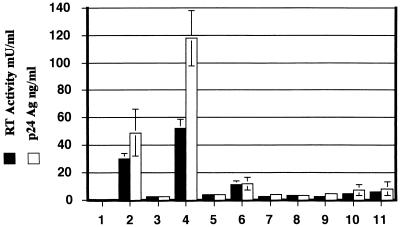
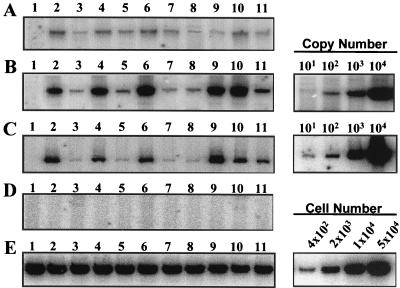
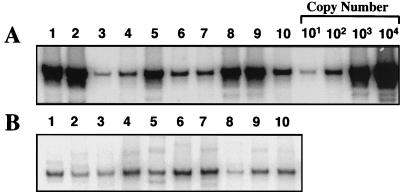
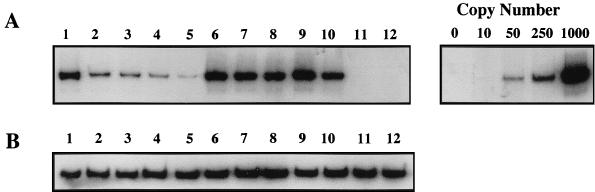
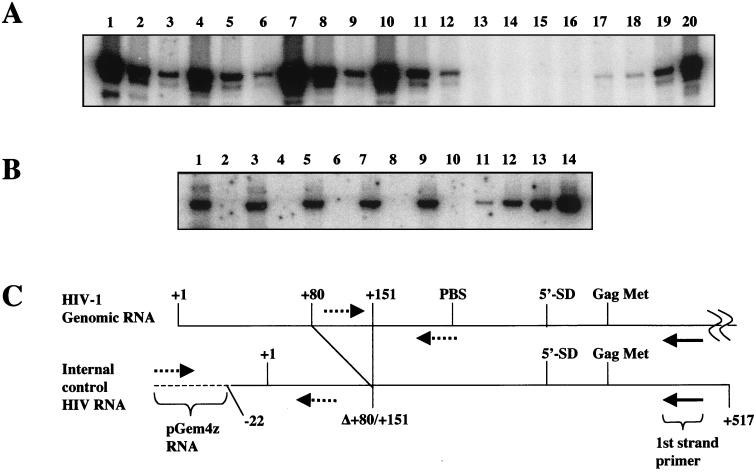
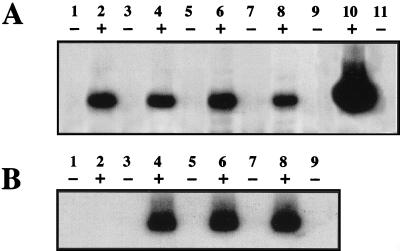
Tat expression is required for efficient human immunodeficiency virus type 1 (HIV-1) reverse transcription. In the present study, we generated a series of 293 cell lines that contained a provirus with a tat gene deletion (Deltatat). Cell lines that contained Deltatat and stably transfected vectors containing either wild-type tat or a number of tat mutants were obtained so that the abilities of these tat genes to stimulate HIV-1 gene expression and reverse transcription could be compared. tat genes with mutations in the amino terminus did not stimulate either viral gene expression or HIV-1 reverse transcription. In contrast, tat mutants in the activation, core, and basic domains of Tat did not stimulate HIV-1 gene expression but markedly stimulated HIV-1 reverse transcription. No differences in the levels of virion genomic RNA or tRNA3Lys were seen in the HIV-1 Deltatat viruses complemented with either mutant or wild-type tat. Finally, overexpression of the Tat-associated kinases CDK7 and CDK9, which are involved in Tat activation of HIV-1 transcription, was not able to complement the reverse transcription defects associated with the lack of a functional tat gene. These results indicate that the mechanism by which tat modulates HIV-1 reverse transcription is distinct from its ability to activate HIV-1 gene expression.
Figures

References
-
- Albini A, Benelli R, Presta M, Rusnati M, Ziche M, Rubartelli A, Paglialunga G, Bussolino F, Noonan D. HIV-tat protein is a heparin-binding angiogenic growth factor. Oncogene. 1996;12:289–297. - PubMed
-
- Albini A, Soldi R, Giunciuglio D, Giraudo E, Benelli R, Primo L, Noonan D, Salio M, Camussi G, Rockl W, Bussolino F. The angiogenesis induced by HIV-1 tat protein is mediated by the Flk-1/KDR receptor on vascular endothelial cells. Nat Med. 1996;2:1371–1375. - PubMed
-
- Baltimore D. RNA-dependent DNA polymerization in virions of RNA tumor viruses. Nature. 1970;226:1209–1211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
