

Review
doi: 10.1128/JB.181.4.1059-1071.1999.
Bacterial adhesins: common themes and variations in architecture and assembly
Affiliations
- PMID: 9973330
- PMCID: PMC93481
- DOI: 10.1128/JB.181.4.1059-1071.1999
Item in Clipboard
Review
Bacterial adhesins: common themes and variations in architecture and assembly
J Bacteriol.
1999 Feb.
No abstract available
Figures
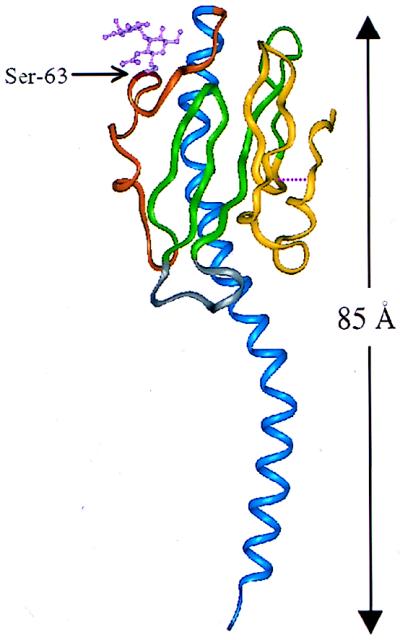
Ribbon representation of pilin from N. gonorrhoeae. Colored regions indicate the secondary structural elements referred to in the text: blue, N-terminal α1-helix (residues 2 to 54); orange, extended disaccharide-bound sugar loop (residues 55 to 77); green, β-hairpins (residues 78 to 93 and 103 to 122); gray, β2-β3 loop (residues 94 to 102); yellow, disulfide bond-containing C-terminal region (residues 121 to 158). Also shown are the disulfide bridge (cysteine residues 121 and 151), signified by a broken line, and Ser-63 with covalently linked disaccharide.
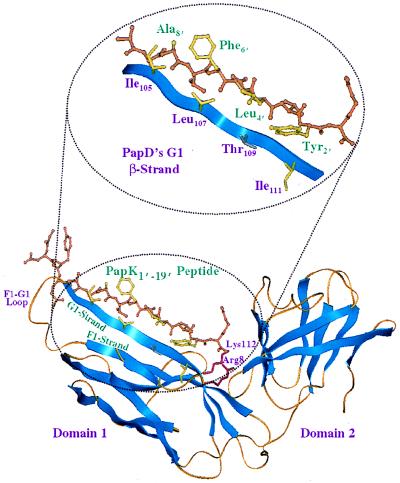
Ribbon representation of the crystal structure of PapD and PapK peptides. Inset provides a magnified view of the PapD and PapK peptide contact interface. Note how the conserved alternating hydrophobic residues of the peptide interdigitate with the residues along PapD’s G1 β-strand.
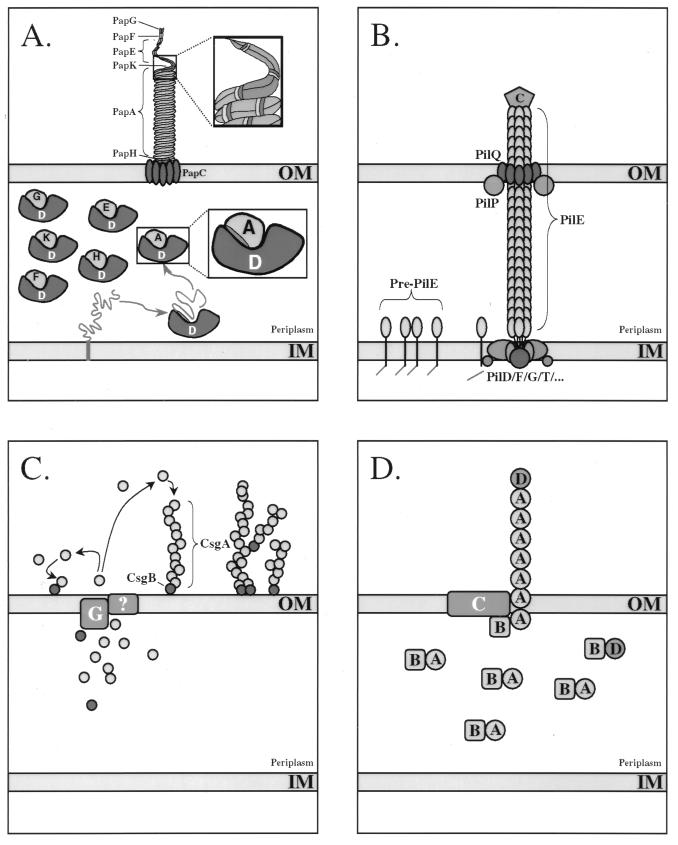
(A) Assembly of P pili from E. coli via the chaperone-usher pathway. Chaperone-mediated extraction of subunits from the inner cytoplasmic membrane (IM) is coupled with their folding into an assembly-competent state. The G1 β-strand of the immunoglobulin-like chaperones, which may serve as a template in the subunit folding pathway, protects nascently folded subunits from premature oligomerization in the periplasmic space by directly capping the newly formed assembly surfaces. These interactive surfaces remain protected by the chaperone until delivery of the preassembly complex to the OM assembly site comprised of the usher. PapG (G), PapD (D), PapE (E), PapK (K), PapA (A), and PapH (H) proteins are shown. (B) Assembly of type IV pilus from N. gonorrhoeae via the general secretion pathway. Prepilin is processed by the PilD signal peptidase, which cleaves the positively charged leader sequence from the N terminus of the pilin subunit. The mature PilE subunit is then assembled by the inner membrane (IM) assembly complex. Translocation of the pilus through the OM is mediated via PilQ, possible with the assistance of other factors such as PilP. The PilC adhesin, which is thought to ultimately be incorporated at the tip of the growing organelle, also appears to be required for translocation through the OM. C, C terminus. (C) Assembly of curli from E. coli via the extracellular nucleation-precipitation pathway. CsgA, the main component of curli, is secreted across the OM. Surface-localized CsgB serves to nucleate CsgA assembly. CsgB is also found distributed along the curli fiber, where it may serve to initiate branching of the fiber. CsgG (G) is an OM-localized lipoprotein that is required for the secretion of CsgA and CsgB, although its function is not known at this time. (D) Assembly of CS1 pili from E. coli via the alternate chaperone pathway. The CooB (B) chaperone forms periplasmic complexes with the main components of the pilus, CooA (A) and CooD (D). It also appears to bind and perhaps stabilize the OM protein CooC (C) in the absence of subunits. CooC may function as an OM channel for passage of the pilin fiber.

Alignment of subunits assembled by FGS chaperones. Amino acid sequence alignment of structural subunits assembled by members of the FGS subfamily of immunoglobulin-like chaperones. Sequences have been grouped into three classes, based upon whether they represent a major or minor subunit and on the morphology of the assembled structures (i.e., thick rods versus thin fimbrillae). Only those residues that are conserved in at least 90% of the sequences within a class or across classes are shaded and coded with the following colors: pink, invariant; yellow, conserved hydrophobic (A, L, V, I, P, M, W, F, C, Y, and G); purple, conserved polar and charged (N, Q, S, T, H, D, E, K, and R). Residues marked by an asterisk appear to be conserved within a given class but not across classes.

Alignment of subunits assembled by FGS chaperones. Amino acid sequence alignment of structural subunits assembled by members of the FGS subfamily of immunoglobulin-like chaperones. Sequences have been grouped into three classes, based upon whether they represent a major or minor subunit and on the morphology of the assembled structures (i.e., thick rods versus thin fimbrillae). Only those residues that are conserved in at least 90% of the sequences within a class or across classes are shaded and coded with the following colors: pink, invariant; yellow, conserved hydrophobic (A, L, V, I, P, M, W, F, C, Y, and G); purple, conserved polar and charged (N, Q, S, T, H, D, E, K, and R). Residues marked by an asterisk appear to be conserved within a given class but not across classes.
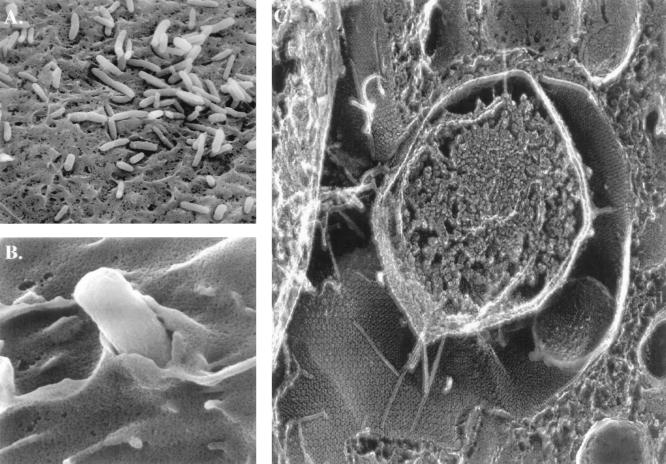
Type 1 pilus-mediated bacterial adherence to the mouse bladder epithelium at 2 h postinfection. (A) Scanning electron micrograph (EM) of bacteria on surface of bladder epithelial cells. The bacteria often appear to be situated in grooves and niches formed by the apical membrane of the superficial cells. (B) Scanning EM of a bacterium being enveloped by the membrane of bladder epithelial cells. (C) High-resolution, freeze-fracture, deep-etch EM of infected bladder epithelia, showing a centrally located bacterium making intimate contact with the luminal surface of the epithelia. Type 1 pili can be seen radiating out from the organisms and spanning the distance between the outer membrane and host cell surface. The FimH adhesin at the tips of these pili mediates contact with the hexagonal uroplakin plaques embedded in the epithelial cell membrane.
References
-
- Adams L M, Simmons C P, Rezmann L, Strugnell R A, Robins-Browne R M. Identification and characterization of a K88- and CS31A-like operon of a rabbit enteropathogenic Escherichia coli strain which encodes fimbriae involved in the colonization of rabbit intestine. Infect Immun. 1997;65:5222–5230. - PMC - PubMed
-
- Alm R A, Mattick J S. Genes involved in the biogenesis and function of type-4 fimbriae in Pseudomonas aeruginosa. Gene. 1997;192:89–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
