

The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast
- PMID: 9990049
- PMCID: PMC15488
- DOI: 10.1073/pnas.96.4.1480
The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast
Abstract
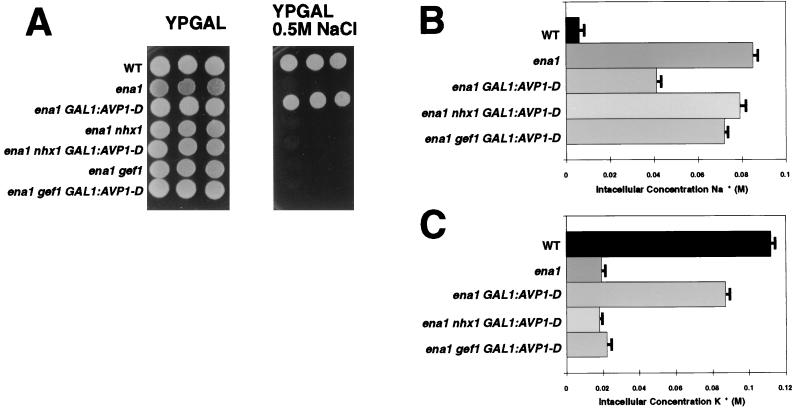
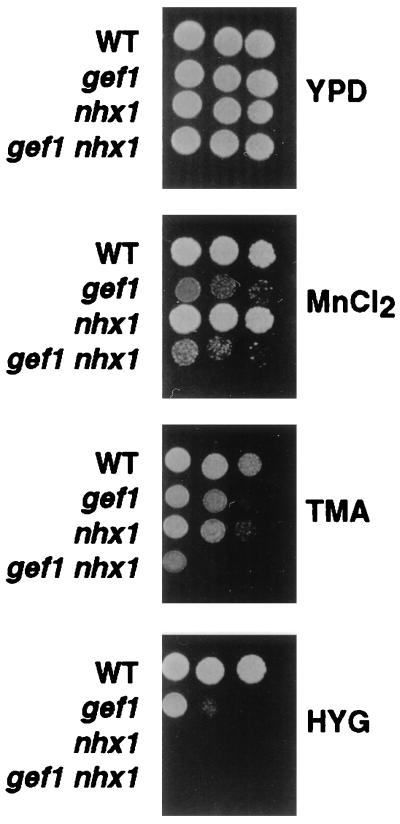
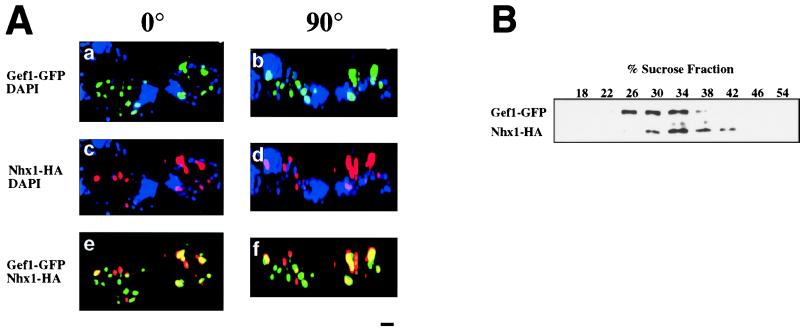
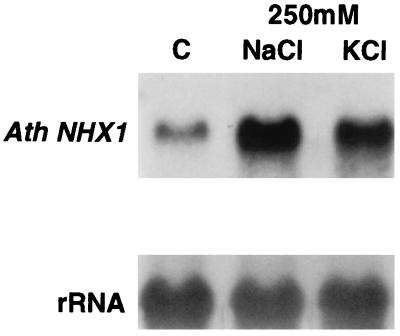
Overexpression of the Arabidopsis thaliana vacuolar H+-pyrophosphatase (AVP1) confers salt tolerance to the salt-sensitive ena1 mutant of Saccharomyces cerevisiae. Suppression of salt sensitivity requires two ion transporters, the Gef1 Cl- channel and the Nhx1 Na+/H+ exchanger. These two proteins colocalize to the prevacuolar compartment of yeast and are thought to be required for optimal acidification of this compartment. Overexpression of AtNHX1, the plant homologue of the yeast Na+/H+ exchanger, suppresses some of the mutant phenotypes of the yeast nhx1 mutant. Moreover, the level of AtNHX1 mRNA in Arabidopsis is increased in the presence of NaCl. The regulation of AtNHX1 by NaCl and the ability of the plant gene to suppress the yeast nhx1 mutant suggest that the mechanism by which cations are detoxified in yeast and plants may be similar.
Figures


References
-
- Haro R, Garciadeblas B, Rodriguez-Navarro A. FEBS Lett. 1991;291:189–191. - PubMed
-
- Rios G, Ferrando A, Serrano R. Yeast. 1997;13:515–528. - PubMed
-
- Prior C, Potier S, Souciet J L, Sychrova H. FEBS Lett. 1996;387:89–93. - PubMed
-
- Dibrov P, Smith J J, Young P G, Fliegel L. FEBS Lett. 1997;405:119–124. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
