

GA-binding protein factors, in concert with the coactivator CREB binding protein/p300, control the induction of the interleukin 16 promoter in T lymphocytes
- PMID: 9990060
- PMCID: PMC15509
- DOI: 10.1073/pnas.96.4.1541
GA-binding protein factors, in concert with the coactivator CREB binding protein/p300, control the induction of the interleukin 16 promoter in T lymphocytes
Abstract
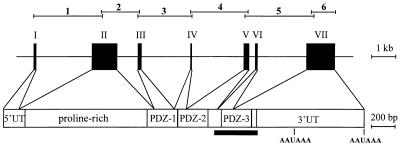
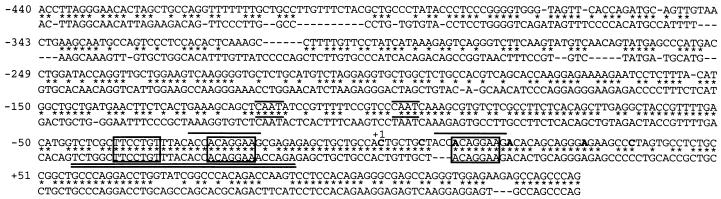
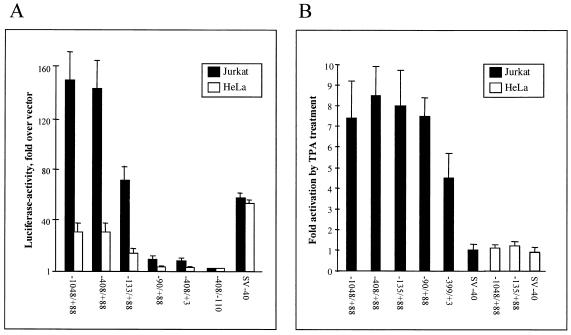
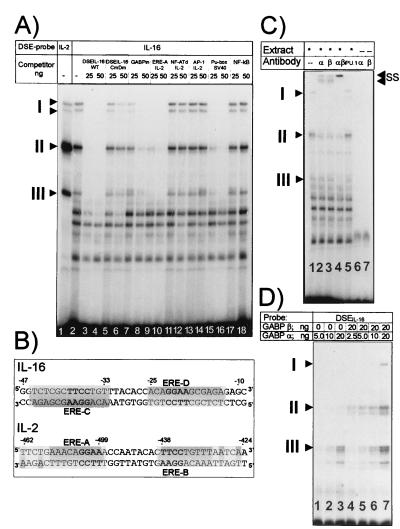
Interleukin 16 (IL-16) is a chemotactic cytokine that binds to the CD4 receptor and affects the activation of T cells and replication of HIV. It is expressed as a large 67-kDa precursor protein (pro-IL-16) in lymphocytes, macrophages, and mast cells, as well as in airway epithelial cells from asthmatics after challenge with allergen. This pro-IL-16 is subsequently processed to the mature cytokine of 13 kDa. To study the expression of IL-16 at the transcriptional level, we cloned the human chromosomal IL-16 gene and analyzed its promoter. The human IL-16 gene consists of seven exons and six introns. The 5' sequences up to nucleotide -120 of the human and murine IL-16 genes share >84% sequence homology and harbor promoter elements for constitutive and inducible transcription in T cells. Although both promoters lack any TATA box, they contain two CAAT box-like motifs and three binding sites of GA-binding protein (GABP) transcription factors. Two of these motifs are part of a highly conserved and inducible dyad symmetry element shown previously to control a remote IL-2 enhancer and the CD18 promoter. In concert with the coactivator CREB binding protein/p300, which interacts with GABPalpha, the binding of GABPalpha and -beta to the dyad symmetry element controls the induction of IL-16 promoter in T cells. Supplementing the data on the processing of pro-IL-16, our results indicate the complexity of IL-16 expression, which is tightly controlled at the transcriptional and posttranslational levels in T lymphocytes.
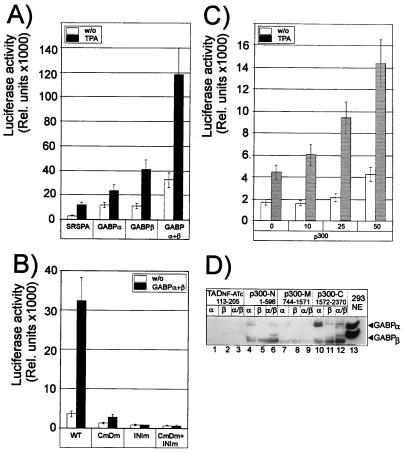
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
