

The PERIANTHIA gene encodes a bZIP protein involved in the determination of floral organ number in Arabidopsis thaliana
- PMID: 9990857
- PMCID: PMC316427
- DOI: 10.1101/gad.13.3.334
The PERIANTHIA gene encodes a bZIP protein involved in the determination of floral organ number in Arabidopsis thaliana
Abstract
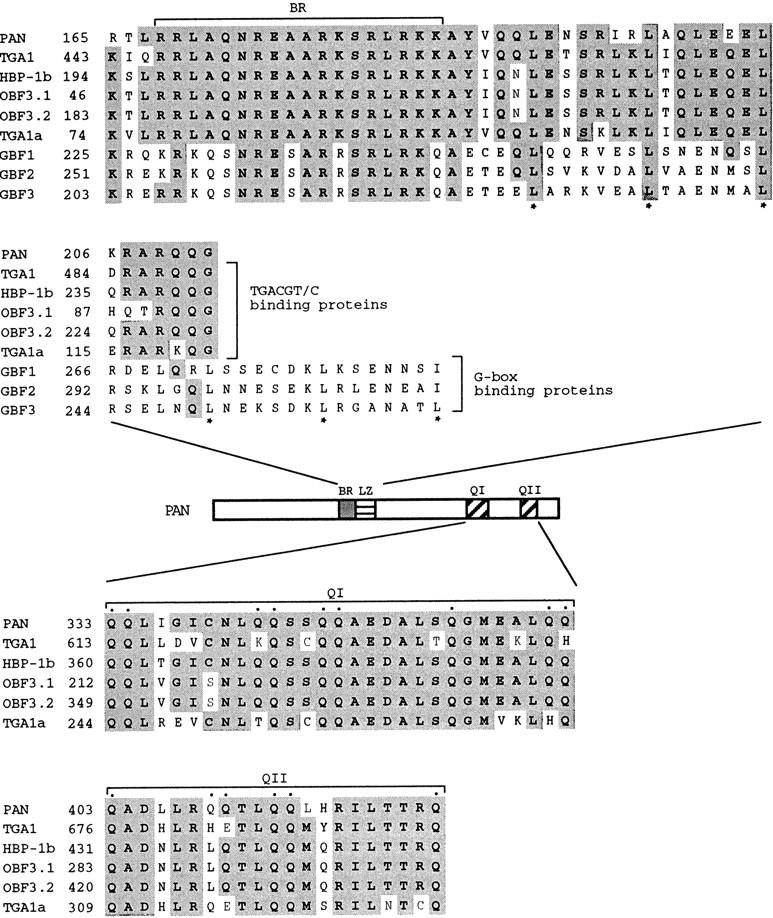
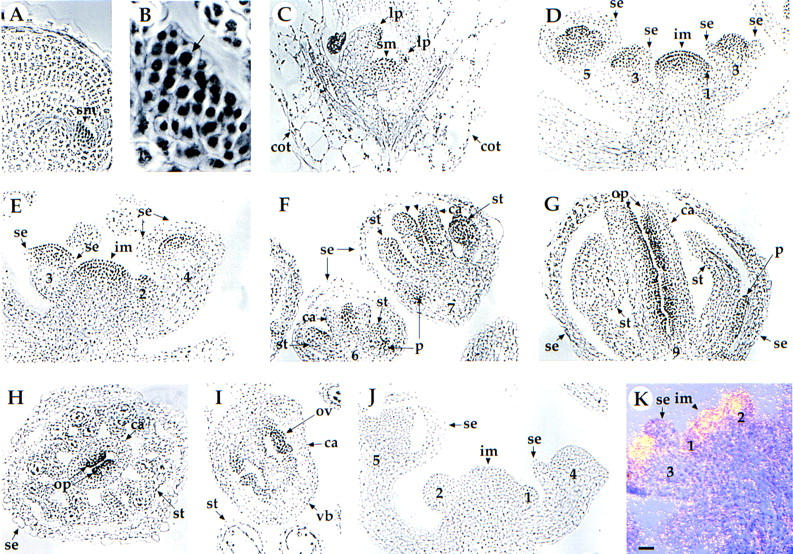
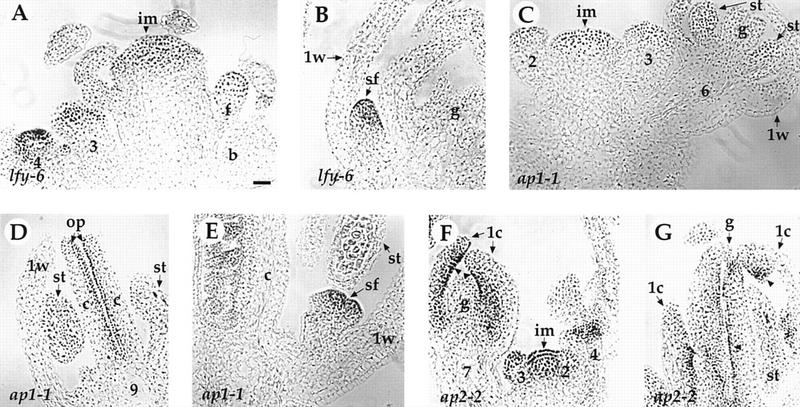
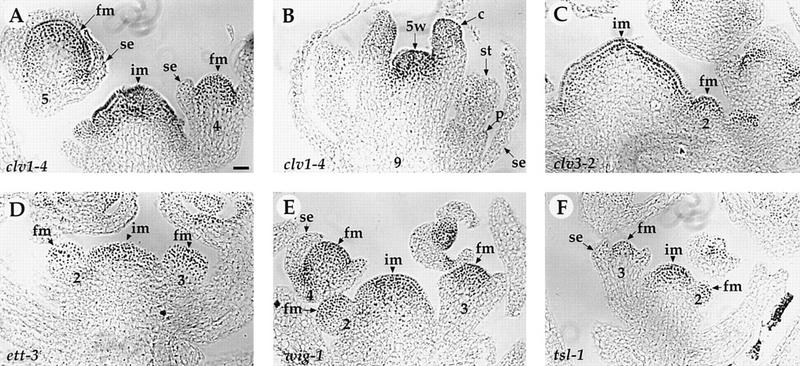
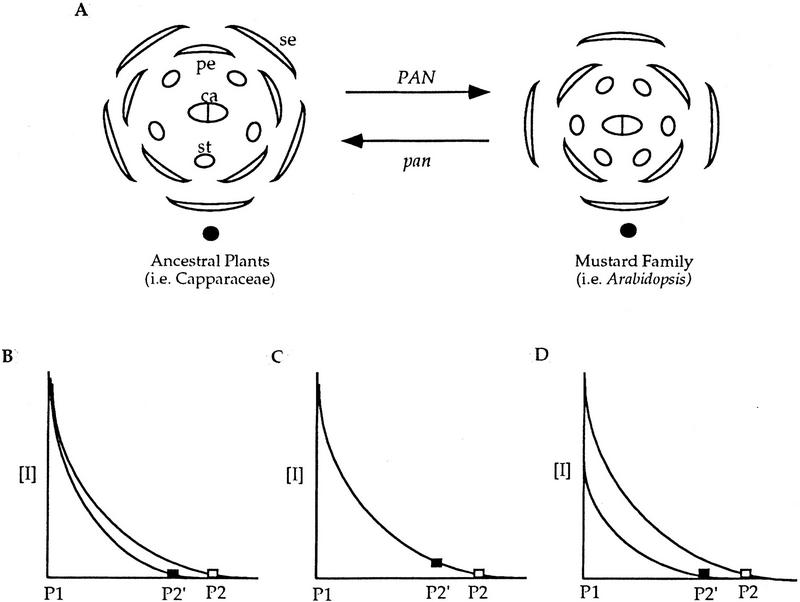
Mutations in the PERIANTHIA (PAN) gene of Arabidopsis thaliana specifically transform flowers from tetramerous to largely pentamerous, which is a characteristic of flowers of ancestral plants. We have cloned the PAN gene and here we show that it encodes a member of the basic region/leucine zipper class of transcription factors. Immunohistochemical analysis shows that the encoded protein is present in the apical meristem, the floral meristem, each whorl of organ primordia, and in ovule primordia during wild-type flower development. PAN expression occurs independently of genes affecting floral meristem identity, floral meristem size, or floral organ number. The near absence of a phenotype in transgenic plants overexpressing PAN and the contrast between the broad expression of PAN and the specificity of its mutant phenotype suggest that its activity may be regulated post-translationally or by the presence of partner proteins. Based on these results and on data reported previously, we propose models for the role of PAN in the evolution of flower pattern in the mustard family.
Figures


References
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. CR Acad Sci. 1993;316:1194–1199.
-
- Bowman JL, editor. Arabidopsis: An atlas of morphology and development. New York, NY: Springer-Verlag; 1994. Flowers: Introduction; pp. 135–145.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases