

Two types of MGDG synthase genes, found widely in both 16:3 and 18:3 plants, differentially mediate galactolipid syntheses in photosynthetic and nonphotosynthetic tissues in Arabidopsis thaliana
- PMID: 11553816
- PMCID: PMC58581
- DOI: 10.1073/pnas.181331498
Two types of MGDG synthase genes, found widely in both 16:3 and 18:3 plants, differentially mediate galactolipid syntheses in photosynthetic and nonphotosynthetic tissues in Arabidopsis thaliana
Abstract
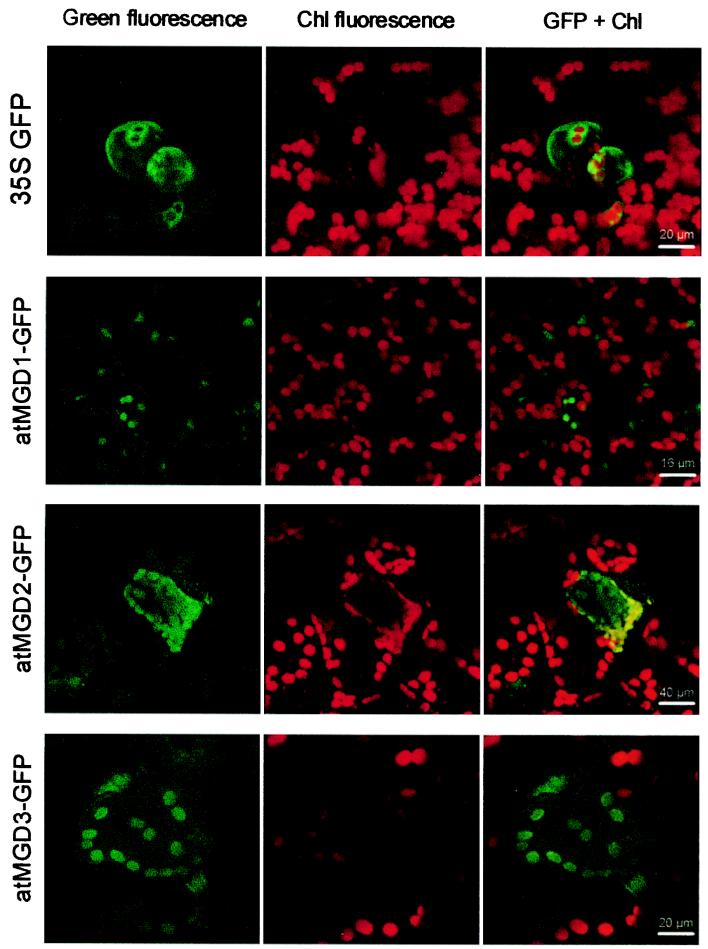
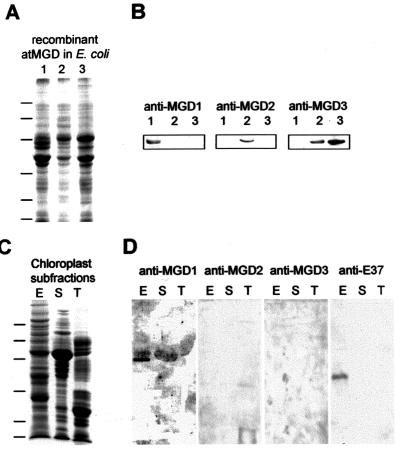
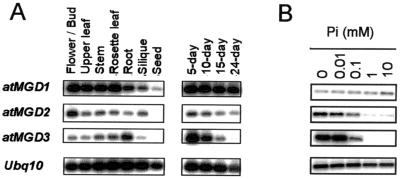
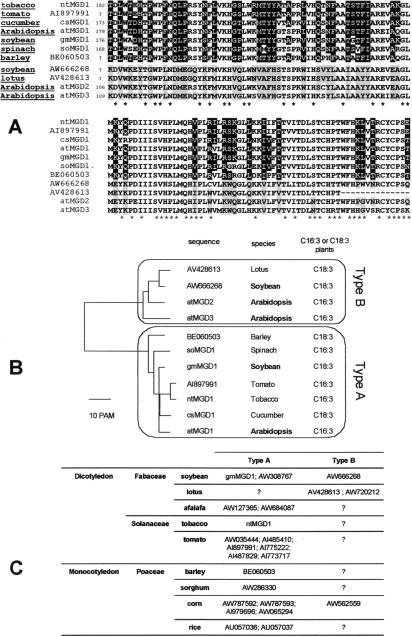
In Arabidopsis, monogalactosyldiacylglycerol (MGDG) is synthesized by a multigenic family of MGDG synthases consisting of two types of enzymes differing in their N-terminal portion: type A (atMGD1) and type B (atMGD2 and atMGD3). The present paper compares type B isoforms with the enzymes of type A that are known to sit in the inner membrane of plastid envelope. The occurrence of types A and B in 16:3 and 18:3 plants shows that both types are not specialized isoforms for the prokaryotic and eukaryotic glycerolipid biosynthetic pathways. Type A atMGD1 gene is abundantly expressed in green tissues and along plant development and encodes the most active enzyme. Its mature polypeptide is immunodetected in the envelope of chloroplasts from Arabidopsis leaves after cleavage of its transit peptide. atMGD1 is therefore likely devoted to the massive production of MGDG required to expand the inner envelope membrane and build up the thylakoids network. Transient expression of green fluorescent protein fusions in Arabidopsis leaves and in vitro import experiments show that type B precursors are targeted to plastids, owing to a different mechanism. Noncanonical addressing peptides, whose processing could not be assessed, are involved in the targeting of type B precursors, possibly to the outer envelope membrane where they might contribute to membrane expansion. Expression of type B enzymes was higher in nongreen tissues, i.e., in inflorescence (atMGD2) and roots (atMGD3), where they conceivably influence the eukaryotic structure prominence in MGDG. In addition, their expression of type B enzymes is enhanced under phosphate deprivation.
Figures
References
-
- Joyard J, Maréchal E, Block M A, Douce R. In: Membranes: Specialized Functions in Plants. Smallwood M, Knox P, Bowles D J, editors. Oxford: BIOS Scientific; 1996. pp. 179–194.
-
- Heinz E. In: Lipids and Lipid Polymers. Tevini M, Lichtenthaler H K, editors. Berlin: Springer; 1977. pp. 102–120.
-
- Block M A, Dorne A-J, Joyard J, Douce R. J Biol Chem. 1983;258:13281–13286. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
