

Diversity in copy number and structure of a silkworm morphogenetic gene as a result of domestication
- PMID: 21242537
- PMCID: PMC3063685
- DOI: 10.1534/genetics.110.124982
Diversity in copy number and structure of a silkworm morphogenetic gene as a result of domestication
Abstract
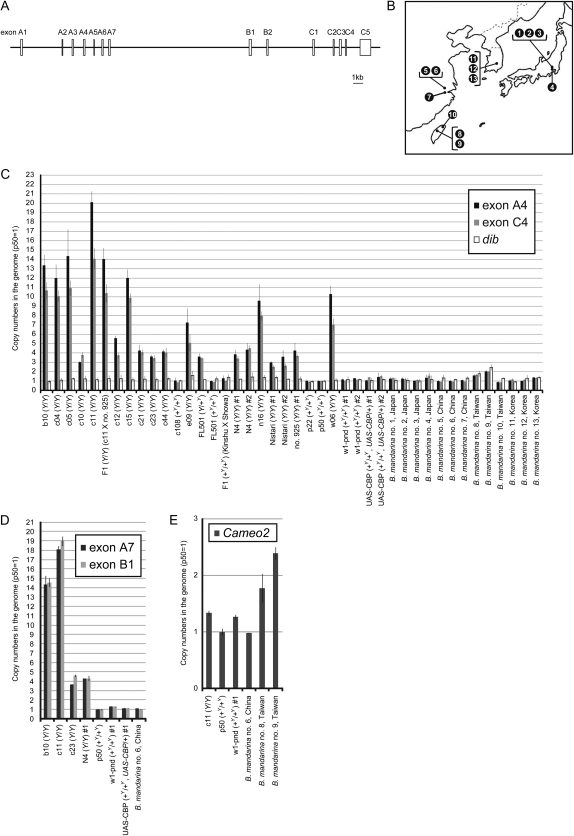
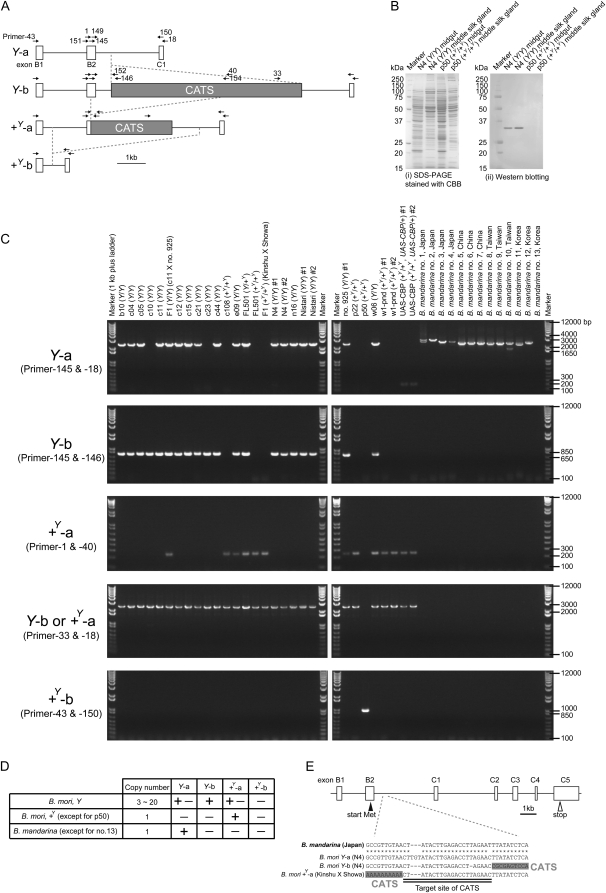
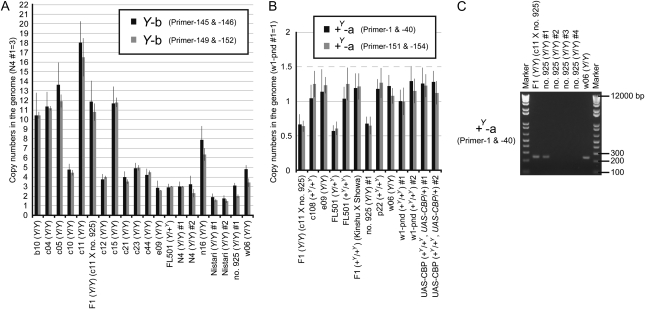
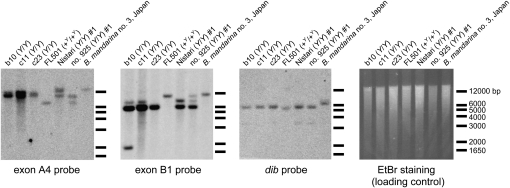
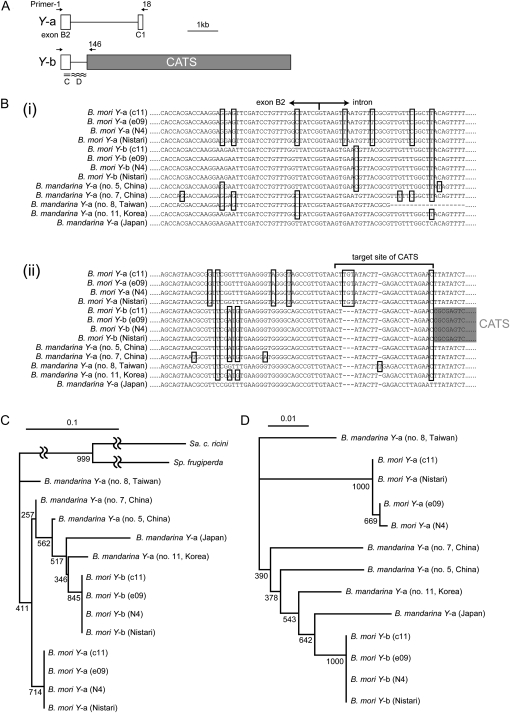
The carotenoid-binding protein (CBP) of the domesticated silkworm, Bombyx mori, a major determinant of cocoon color, is likely to have been substantially influenced by domestication of this species. We analyzed the structure of the CBP gene in multiple strains of B. mori, in multiple individuals of the wild silkworm, B. mandarina (the putative wild ancestor of B. mori), and in a number of other lepidopterans. We found the CBP gene copy number in genomic DNA to vary widely among B. mori strains, ranging from 1 to 20. The copies of CBP are of several types, based on the presence of a retrotransposon or partial deletion of the coding sequence. In contrast to B. mori, B. mandarina was found to possess a single copy of CBP without the retrotransposon insertion, regardless of habitat. Several other lepidopterans were found to contain sequences homologous to CBP, revealing that this gene is evolutionarily conserved in the lepidopteran lineage. Thus, domestication can generate significant diversity of gene copy number and structure over a relatively short evolutionary time.
© 2011 by the Genetics Society of America
Figures


References
-
- Alpy, F., and C. Tomasetto, 2006. MLN64 and MENTHO, two mediators of endosomal cholesterol transport. Biochem. Soc. Trans. 34 343–345. - PubMed
-
- Andersson, D. I., and D. Hughes, 2009. Gene amplification and adaptive evolution in bacteria. Annu. Rev. Genet. 43 167–195. - PubMed
-
- Arunkumar, K. P., M. Metta and J. Nagaraju, 2006. Molecular phylogeny of silkmoths reveals the origin of domesticated silkmoth, Bombyx mori from Chinese Bombyx mandarina and paternal inheritance of Antheraea proylei mitochondrial DNA. Mol. Phylogenet. Evol. 40 419–427. - PubMed
-
- Banno, Y., T. Shimada, Z. Kajiura and H. Sezutsu, 2010. The silkworm-an attractive BioResource supplied by Japan. Exp. Anim. 59 139–146. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
