

doi: 10.1371/journal.pgen.0030032.
Elevated rates of sister chromatid exchange at chromosome ends
Affiliations
- PMID: 17319749
- PMCID: PMC1802831
- DOI: 10.1371/journal.pgen.0030032
Item in Clipboard
Elevated rates of sister chromatid exchange at chromosome ends
PLoS Genet.
.
Abstract
Chromosome ends are known hotspots of meiotic recombination and double-strand breaks. We monitored mitotic sister chromatid exchange (SCE) in telomeres and subtelomeres and found that 17% of all SCE occurs in the terminal 0.1% of the chromosome. Telomeres and subtelomeres are significantly enriched for SCEs, exhibiting rates of SCE per basepair that are at least 1,600 and 160 times greater, respectively, than elsewhere in the genome.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
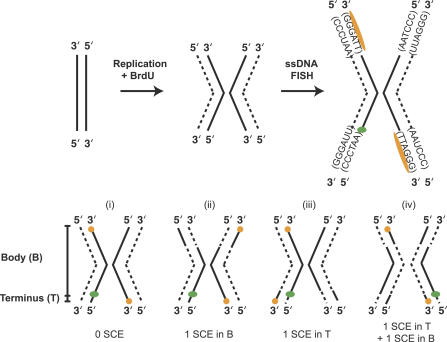
Cells were synchronized at the G1/S boundary and allowed to replicate in the presence of BrdU. Replicated chromosomes have chromatids with one BrdU-incorporated DNA strand (dashed line) and one original DNA strand (solid line). The BrdU-incorporated strand is digested so that it is unavailable for hybridization. The single-stranded telomere probe (orange) and one single-stranded internal subtelomeric probe (green) hybridize to the non-BrdU-labeled strand. Each of four internal probes was cohybridized with the telomeric probe in a separate experiment. Telomere repeats, (TTAGGG)n, are oriented 5′ to 3′, so the orange telomeric PNA probe, (CCCTAA)3, hybridizes to one chromatid on each chromosome end. Fully degraded and hybridized chromosomes may exhibit one of four probe configurations. (i) If no SCE has occurred during or subsequent to replication, telomeres are oriented in a diagonal (trans) pattern, and the internal probe, because of its 5′–3′ orientation, hybridizes to the chromatid opposite the nearest telomere signal. (ii) If one SCE, or an odd number of SCEs, occur(s) in the body of the chromosome (B), as indicated by switched dashed and solid lines, the telomere probes hybridize to both ends of the same chromatid (in a cis pattern), but the internal probe remains at the same position relative to the closest telomere signal (i.e., on the opposite chromatid). (iii) If one or an odd number of SCEs occurs in the terminal interval (T) between the subtelomeric internal probe and the telomere, all probe signals will lie on the same chromatid. (iv) If one SCE occurs in the body and one occurs in the terminal interval of the chromosome, the telomere probe signals will appear in their original diagonal trans configuration, but the internal subtelomere probe will shift to the opposite chromatid than in (i) and end up on the same chromatid as the nearby telomere signal.
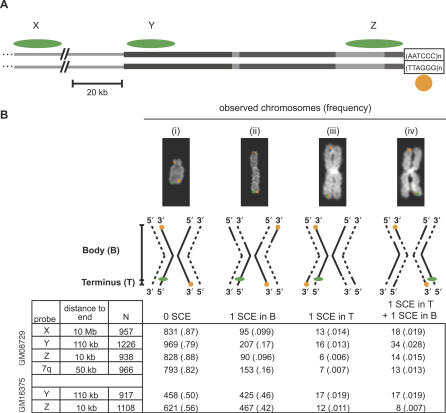
(A) Single-stranded CO-FISH probes X, Y, and Z, shown as green ovals, hybridize 10 Mb, 110 kb, and 10 kb from the end of chromosomes, respectively. Y and Z probes hybridize to duplicated subtelomeric sequences, shown as grey rectangles. The orange probe indicated by the circle hybridizes to the telomere-repeat sequence (TTAGGG)n at the ends of all chromosomes. The amount of SCE occurring between the orange telomeric signal and each green probe signal was measured in separate experiments. The scale bar indicates 20 kb. (B) In each experiment, four different CO-FISH configurations are possible in fully processed and hybridized chromosomes, shown as examples and diagrams as in Figure 1. The number of chromosomes (and relative frequency) observed in each configuration is shown below. SCE events within the telomere that split the telomere probe signal between two chromatids at the same end were counted separately and not included in our estimates of terminal SCE rates (Text S1). GM08729 is a normal lymphoblastoid cell line, and GM16375 is a lymphoblastoid cell line derived from a patient with Bloom syndrome.
References
-
- Kong A, Gudbjartsson DF, Sainz J, Jonsdottir GM, Gudjonsson SA, et al. A high-resolution recombination map of the human genome. Nat Genet. 2002;31:241–247. - PubMed
-
- d'Adda di Fagagna F, Reaper PM, Clay-Farrace L, Fiegler H, Carr P, et al. A DNA damage checkpoint response in telomere-initiated senescence. Nature. 2003;426:194–198. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
