

Evolution of human-specific neural SRGAP2 genes by incomplete segmental duplication
- PMID: 22559943
- PMCID: PMC3365555
- DOI: 10.1016/j.cell.2012.03.033
Evolution of human-specific neural SRGAP2 genes by incomplete segmental duplication
Abstract
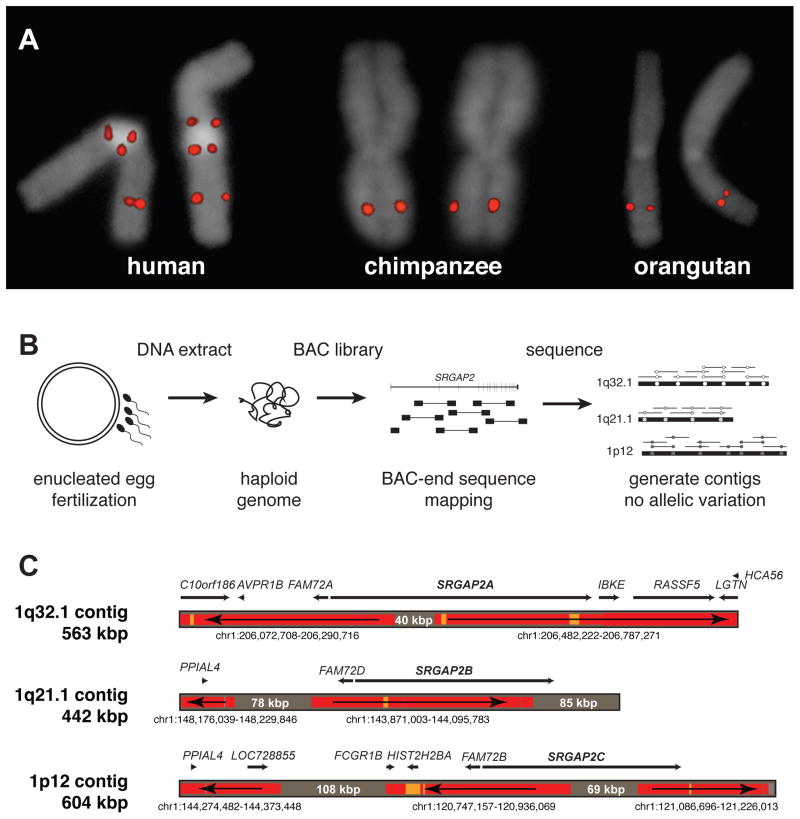
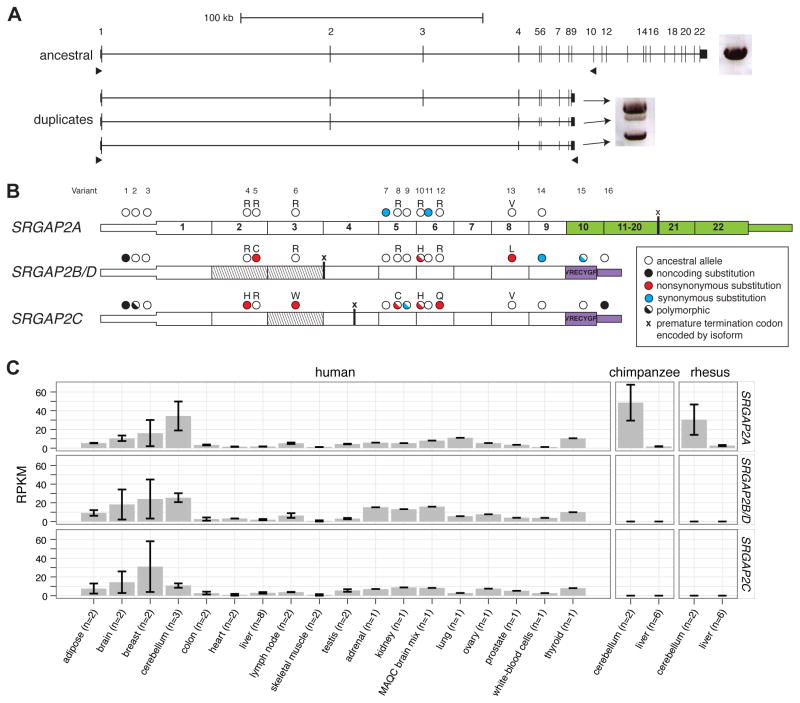
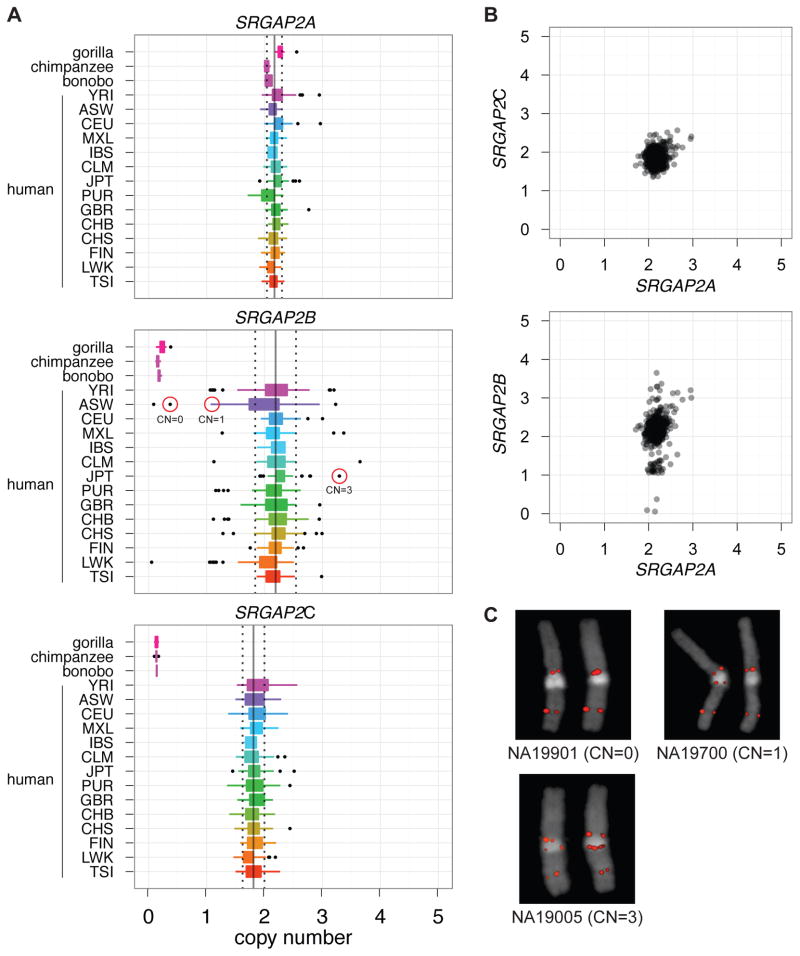
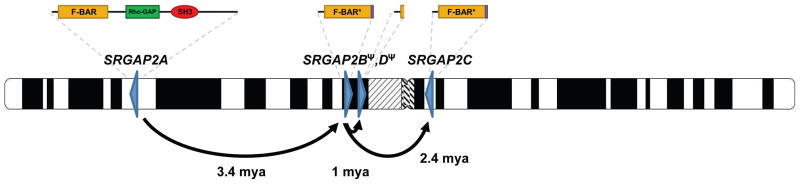
Gene duplication is an important source of phenotypic change and adaptive evolution. We leverage a haploid hydatidiform mole to identify highly identical sequences missing from the reference genome, confirming that the cortical development gene Slit-Robo Rho GTPase-activating protein 2 (SRGAP2) duplicated three times exclusively in humans. We show that the promoter and first nine exons of SRGAP2 duplicated from 1q32.1 (SRGAP2A) to 1q21.1 (SRGAP2B) ∼3.4 million years ago (mya). Two larger duplications later copied SRGAP2B to chromosome 1p12 (SRGAP2C) and to proximal 1q21.1 (SRGAP2D) ∼2.4 and ∼1 mya, respectively. Sequence and expression analyses show that SRGAP2C is the most likely duplicate to encode a functional protein and is among the most fixed human-specific duplicate genes. Our data suggest a mechanism where incomplete duplication created a novel gene function-antagonizing parental SRGAP2 function-immediately "at birth" 2-3 mya, which is a time corresponding to the transition from Australopithecus to Homo and the beginning of neocortex expansion.
Copyright © 2012 Elsevier Inc. All rights reserved.
Conflict of interest statement
J.A.R. and L.S. are employees of Signature Genomic Laboratories, a subsidiary of PerkinElmer, Inc. E.E.E. is on the scientific advisory boards for Pacific Biosciences, Inc. and SynapDx Corp.
Figures

Comment in
-
Sibling rivalry among paralogs promotes evolution of the human brain.Cell. 2012 May 11;149(4):737-9. doi: 10.1016/j.cell.2012.04.020. Cell. 2012. PMID: 22579279 Free PMC article.
-
Genome evolution: Functional antagonism and human brain evolution.Nat Rev Genet. 2012 Jul;13(7):452. doi: 10.1038/nrg3266. Nat Rev Genet. 2012. PMID: 22673161 No abstract available.
References
-
- Bailey JA, Gu Z, Clark RA, Reinert K, Samonte RV, Schwartz S, Adams MD, Myers EW, Li PW, Eichler EE. Recent segmental duplications in the human genome. Science. 2002;297:1003–1007. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
