

Phylogenetic affinity of a wide, vacuolate, nitrate-accumulating Beggiatoa sp. from Monterey Canyon, California, with Thioploca spp
- PMID: 9872789
- PMCID: PMC91012
- DOI: 10.1128/AEM.65.1.270-277.1999
Phylogenetic affinity of a wide, vacuolate, nitrate-accumulating Beggiatoa sp. from Monterey Canyon, California, with Thioploca spp
Abstract
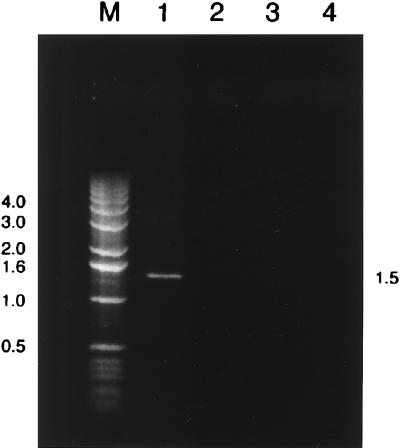
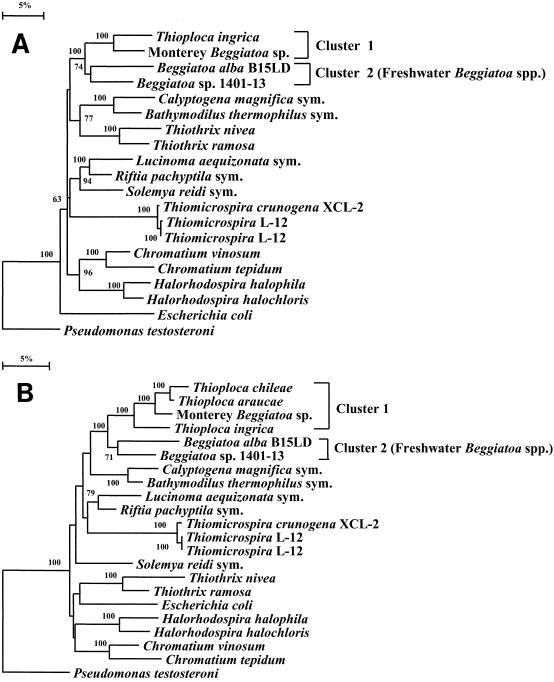
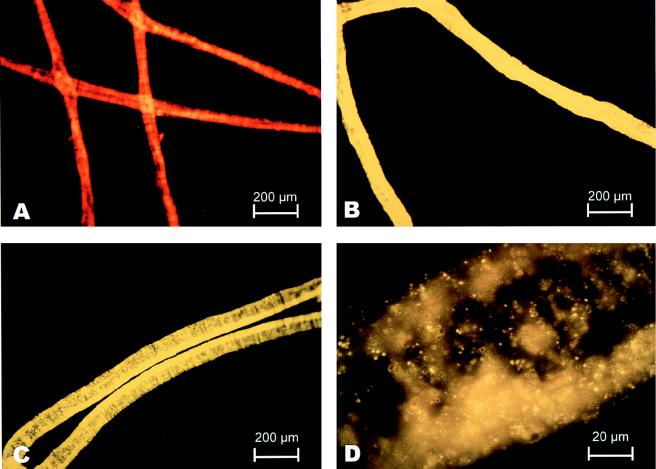
Environmentally dominant members of the genus Beggiatoa and Thioploca spp. are united by unique morphological and physiological adaptations (S. C. McHatton, J. P. Barry, H. W. Jannasch, and D. C. Nelson, Appl. Environ. Microbiol. 62:954-958, 1996). These adaptations include the presence of very wide filaments (width, 12 to 160 microm), the presence of a central vacuole comprising roughly 80% of the cellular biovolume, and the capacity to internally concentrate nitrate at levels ranging from 150 to 500 mM. Until recently, the genera Beggiatoa and Thioploca were recognized and differentiated on the basis of morphology alone; they were distinguished by the fact that numerous Thioploca filaments are contained within a common polysaccharide sheath, while Beggiatoa filaments occur singly. Vacuolate Beggiatoa or Thioploca spp. can dominate a variety of marine sediments, seeps, and vents, and it has been proposed (H. Fossing, V. A. Gallardo, B. B. Jorgensen, M. Huttel, L. P. Nielsen, H. Schulz, D. E. Canfield, S. Forster, R. N. Glud, J. K. Gundersen, J. Kuver, N. B. Ramsing, A. Teske, B. Thamdrup, and O. Ulloa, Nature [London] 374:713-715, 1995) that members of the genus Thioploca are responsible for a significant portion of total marine denitrification. In order to investigate the phylogeny of an environmentally dominant Beggiatoa sp., we analyzed complete 16S rRNA gene sequence data obtained from a natural population found in Monterey Canyon cold seeps. Restriction fragment length polymorphism analysis of a clone library revealed a dominant clone, which gave rise to a putative Monterey Beggiatoa 16S rRNA sequence. Fluorescent in situ hybridization with a sequence-specific probe confirmed that this sequence originated from wide Beggiatoa filaments (width, 65 to 85 microm). A phylogenetic tree based on evolutionary distances indicated that the Monterey Beggiatoa sp. falls in the gamma subdivision of the class Proteobacteria and is most closely related to the genus Thioploca. This vacuolate Beggiatoa-Thioploca cluster and a more distantly related freshwater Beggiatoa species cluster form a distinct phylogenetic group.
Figures


References
-
- Barry J P, Greene H G, Orange D L, Baxter C H, Robinson B H, Kochevar R E, Nybakken J W, Reed D L, McHugh C M. Biologic and geologic characteristics of cold seeps in Monterey Bay, California. Deep Sea Res Part A Oceanogr Res Pap. 1996;43:1739–1762.
-
- Buck, K. Personal communication.
-
- Felenstein J. PHYLIP—phylogeny inference package. Cladistics. 1989;5:164–166.
-
- Fossing H, Gallardo V A, Jorgensen B B, Huttel M, Nielson L P, Schulz H, Canfield D E, Forster S, Glud R N, Gundersen J K, Kuver J, Ramsing N B, Teske A, Thamdrup B, Ulloa O. Concentration and transport of nitrate by the mat-forming sulphur bacterium Thioploca. Nature (London) 1995;374:713–716.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
