

Surface cytotoxic T lymphocyte-associated antigen 4 partitions within lipid rafts and relocates to the immunological synapse under conditions of inhibition of T cell activation
- PMID: 12021313
- PMCID: PMC2193751
- DOI: 10.1084/jem.20011868
Surface cytotoxic T lymphocyte-associated antigen 4 partitions within lipid rafts and relocates to the immunological synapse under conditions of inhibition of T cell activation
Abstract
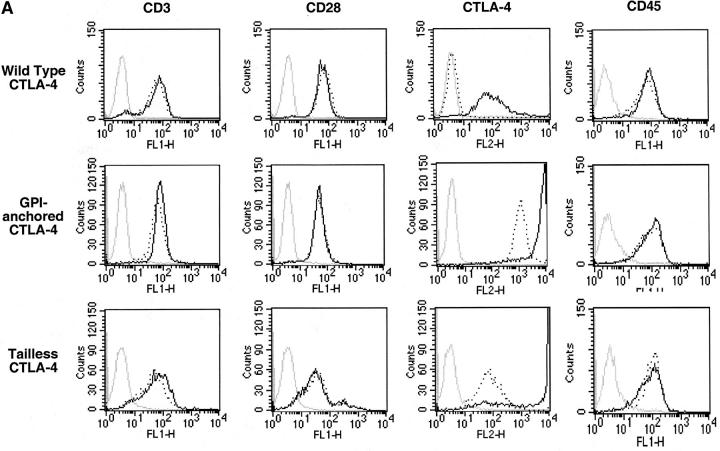
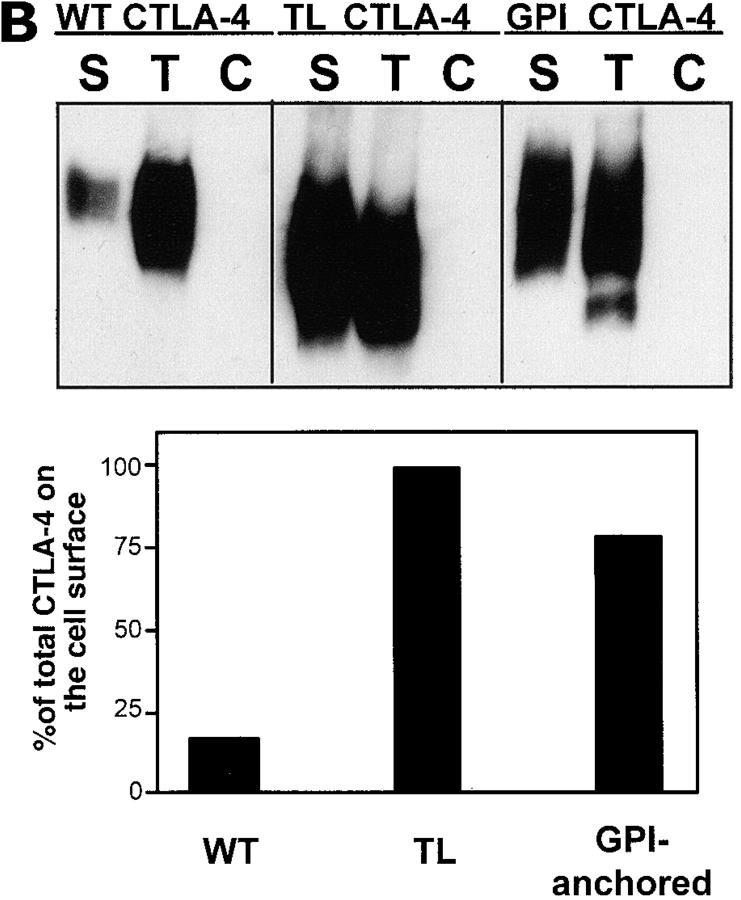
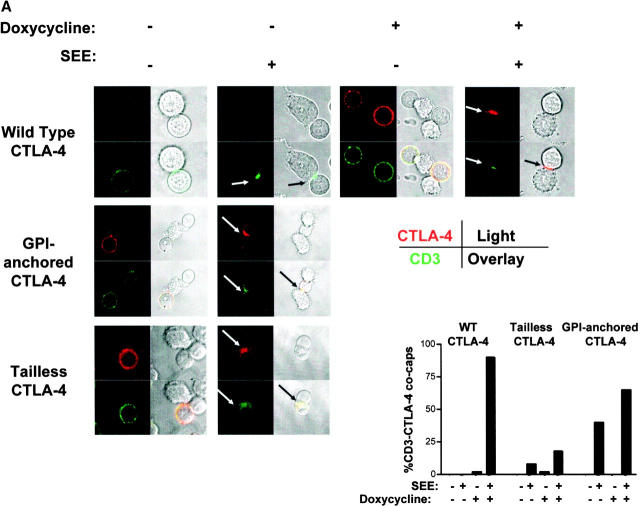
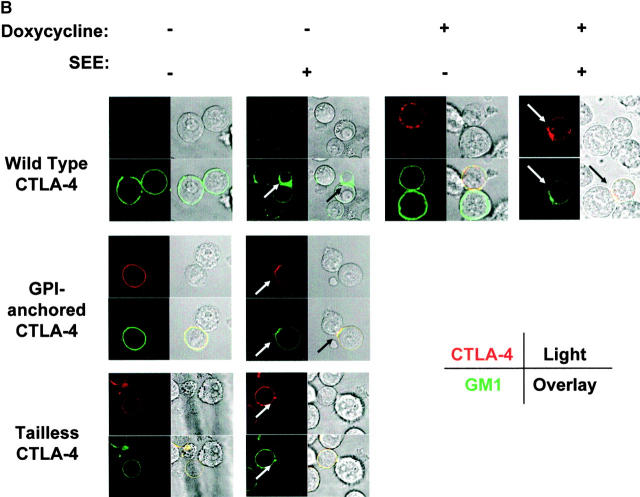
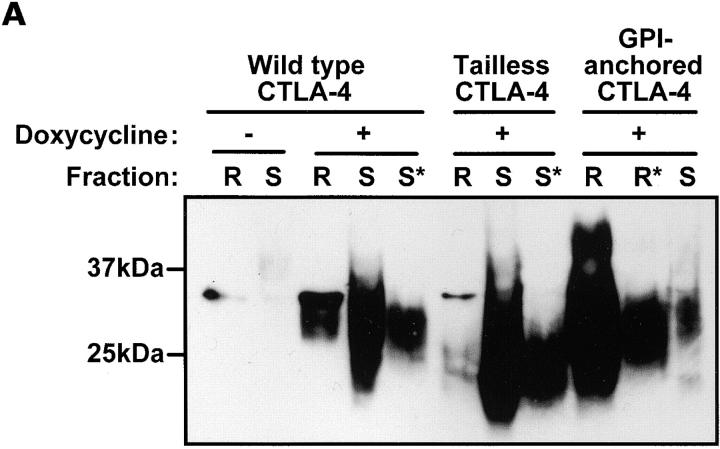
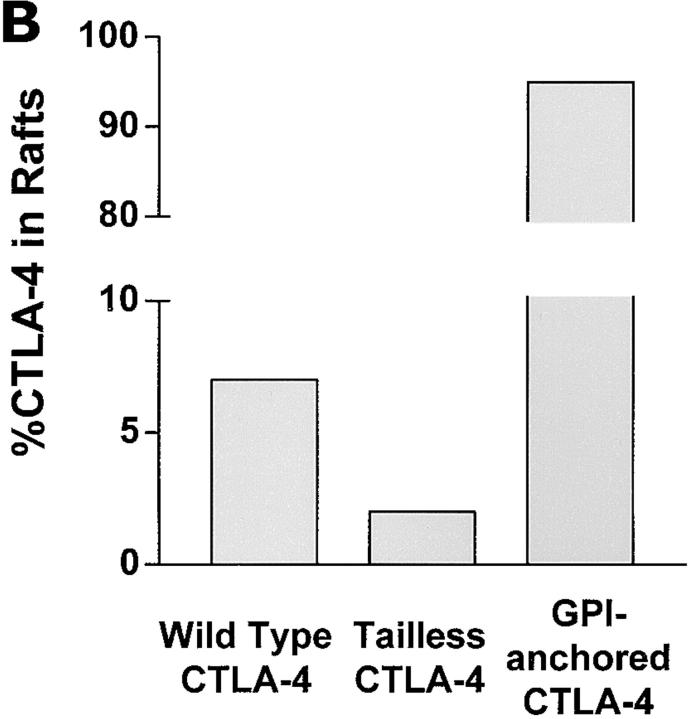
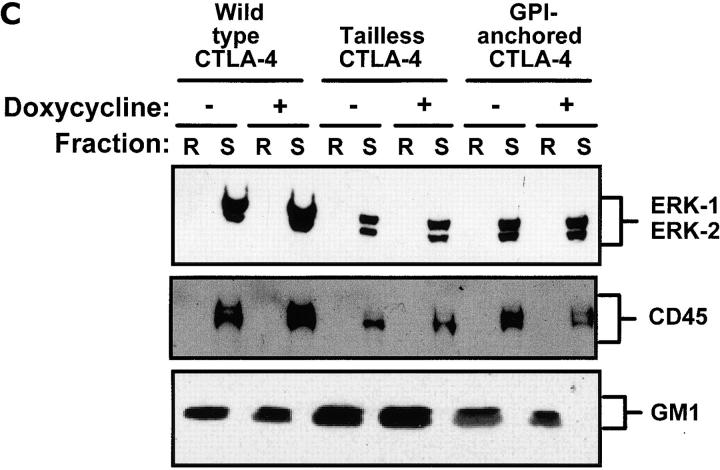
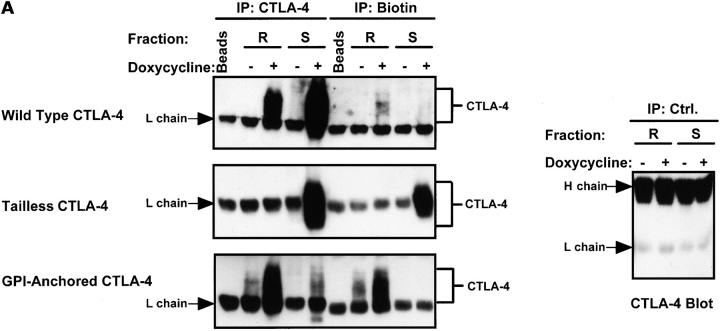
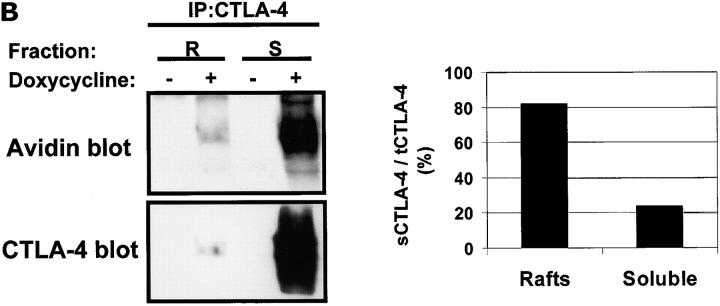
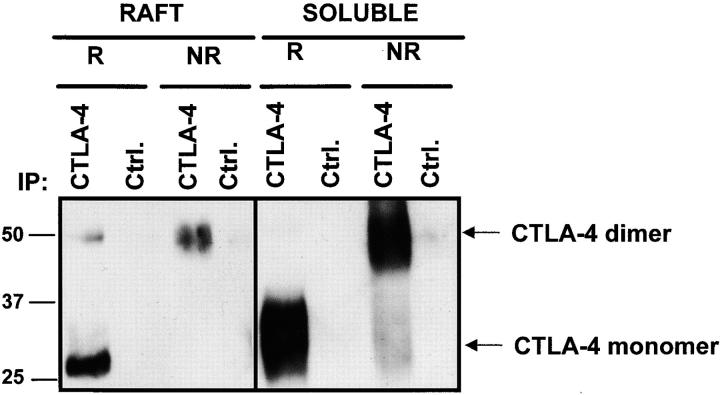
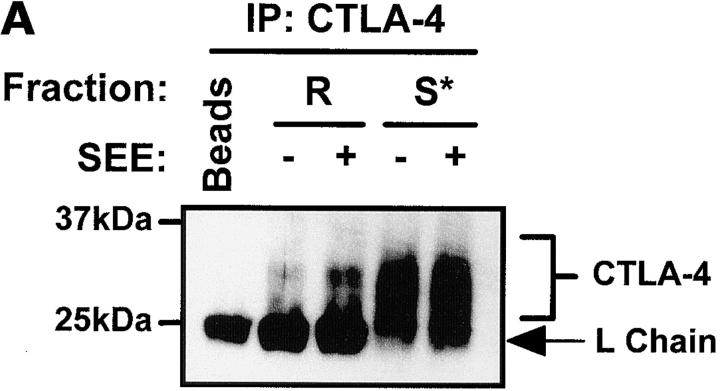
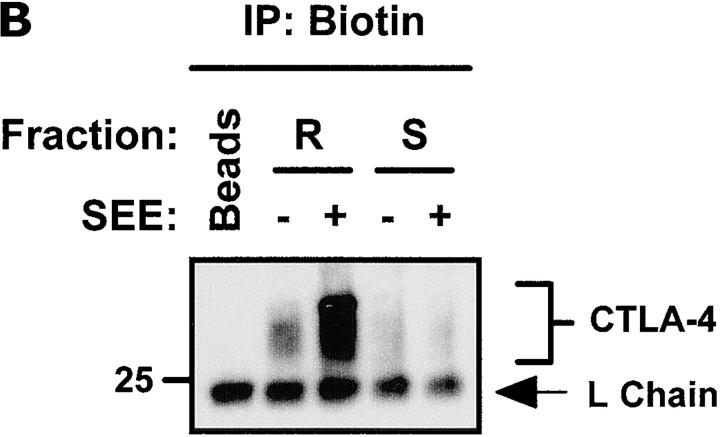
T cell activation through the T cell receptor (TCR) involves partitioning of receptors into discrete membrane compartments known as lipid rafts, and the formation of an immunological synapse (IS) between the T cell and antigen-presenting cell (APC). Compartmentalization of negative regulators of T cell activation such as cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) is unknown. Recent crystal structures of B7-ligated CTLA-4 suggest that it may form lattices within the IS which could explain the mechanism of action of this molecule. Here, we show that after T cell stimulation, CTLA-4 coclusters with the TCR and the lipid raft ganglioside GM1 within the IS. Using subcellular fractionation, we show that most lipid raft-associated CTLA-4 is on the T cell surface. Such compartmentalization is dependent on the cytoplasmic tail of CTLA-4 and can be forced with a glycosylphosphatidylinositol-anchor in CTLA-4. The level of CTLA-4 within lipid rafts increases under conditions of APC-dependent TCR-CTLA-4 coligation and T cell inactivation. However, raft localization, although necessary for inhibition of T cell activation, is not sufficient for CTLA-4-mediated negative signaling. These data demonstrate that CTLA-4 within lipid rafts migrates to the IS where it can potentially form lattice structures and inhibit T cell activation.
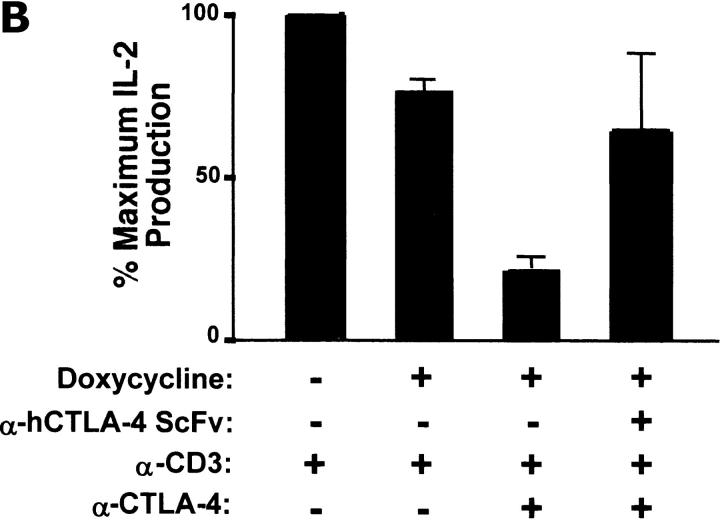
Figures
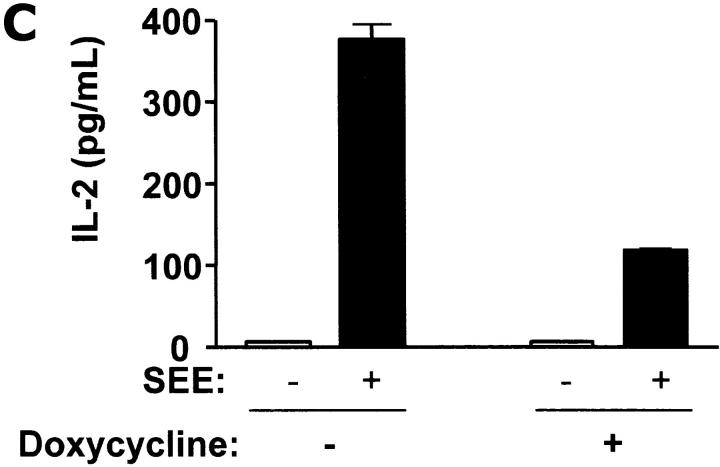
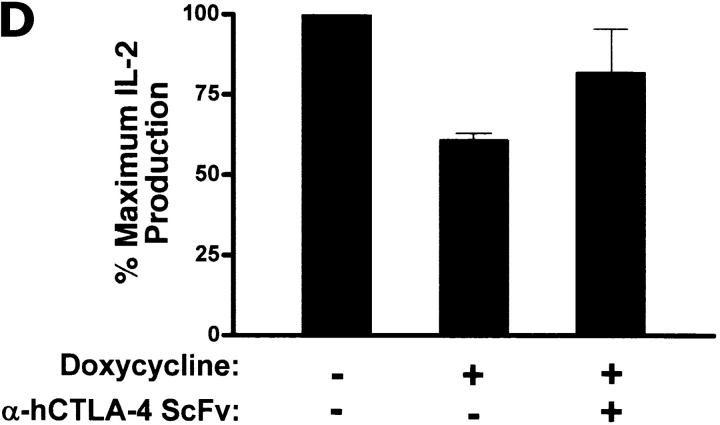
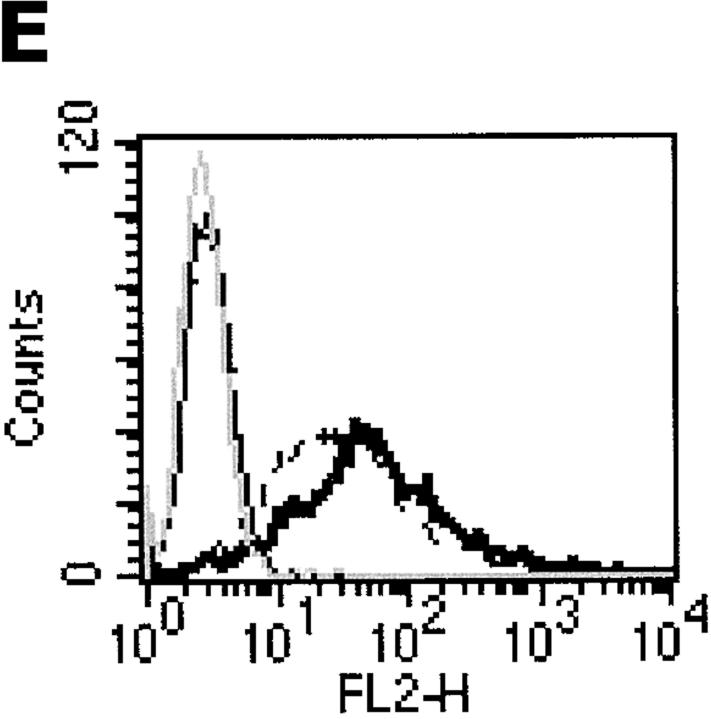
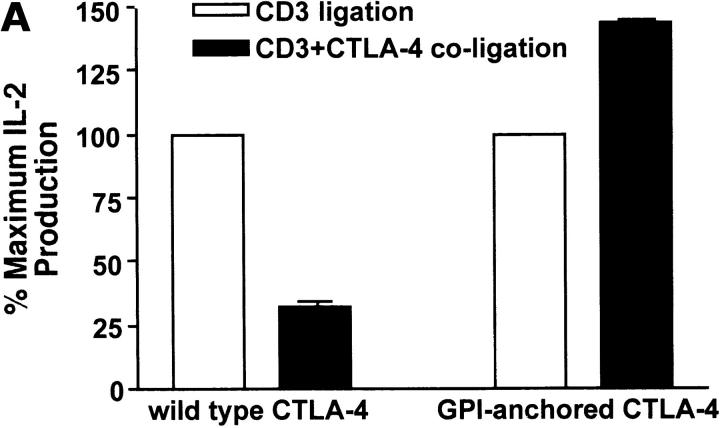
References
-
- Bromley, S.K., W.R. Burack, K.G. Johnson, K. Somersalo, T.N. Sims, C. Sumen, M.M. Davis, A.S. Shaw, P.M. Allen, and M.L. Dustin. 2001. The immunological synapse. Annu. Rev. Immunol. 19:375–396. - PubMed
-
- Monks, C.R., H. Kupfer, I. Tamir, A. Barlow, and A. Kupfer. 1997. Selective modulation of protein kinase C-theta during T-cell activation. Nature. 385:83–86. - PubMed
-
- Monks, C.R., B.A. Freiberg, H. Kupfer, N. Sciaky, and A. Kupfer. 1998. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 395:82–86. - PubMed
-
- Delon, J., K. Kaibuchi, and R.N. Germain. 2001. Exclusion of CD43 from the immunological synapse is mediated by phosphorylation-regulated relocation of the cytoskeletal adaptor moesin. Immunity. 15:691–701. - PubMed
-
- Shaw, A.S. 2001. FERMing up the synapse. Immunity. 15:683–686. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
