

Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. II. Accumulation of plant mRNAs in response to insect-derived cues
- PMID: 11161027
- PMCID: PMC64871
- DOI: 10.1104/pp.125.2.701
Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. II. Accumulation of plant mRNAs in response to insect-derived cues
Abstract
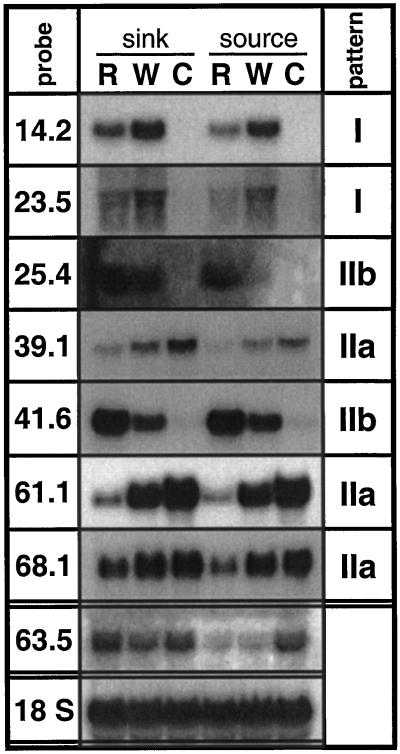
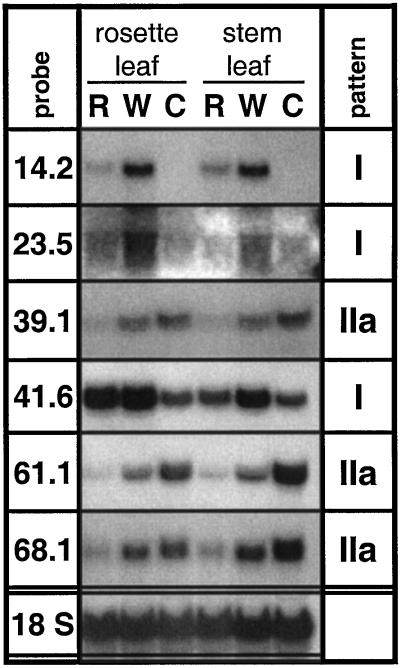
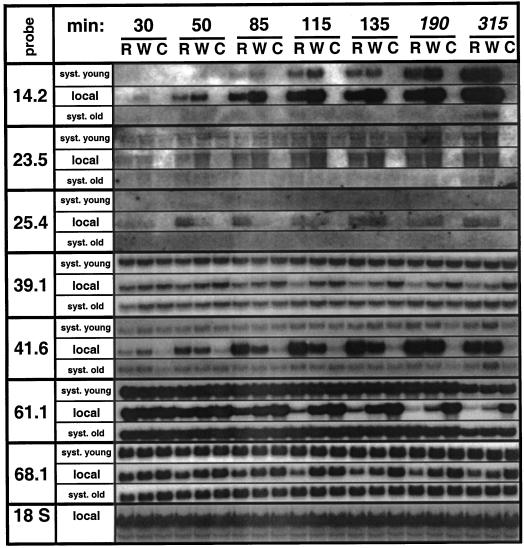
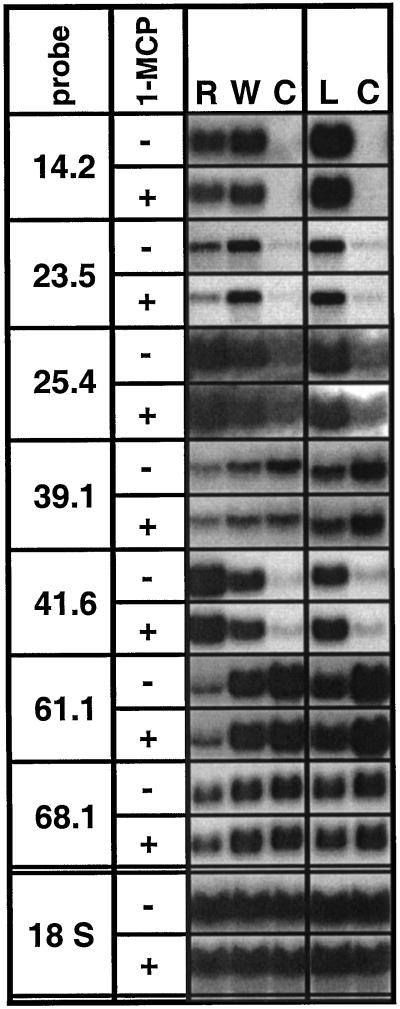
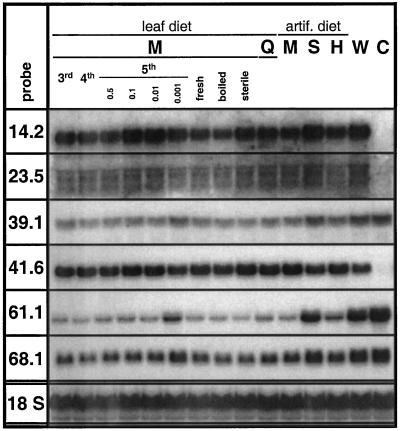
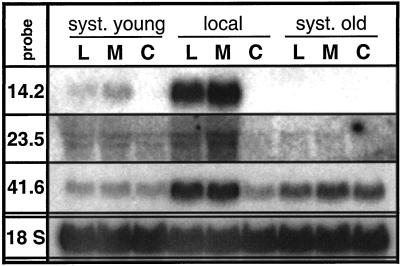
The transcriptional changes in Nicotiana attenuata Torr. ex Wats. elicited by attack from Manduca sexta larvae were previously characterized by mRNA differential display (D. Hermsmeier, U. Schittko, I.T. Baldwin [2001] Plant Physiol 125: 683-700). Because herbivore attack causes wounding, we disentangled wound-induced changes from those elicited by M. sexta oral secretions and regurgitant (R) with a northern analysis of a subset of the differentially expressed transcripts encoding threonine deaminase, pathogen-induced oxygenase, a photosystem II light-harvesting protein, a retrotransposon homolog, and three unknown genes. R extensively modified wound-induced responses by suppressing wound-induced transcripts (type I) or amplifying the wound-induced response (type II) further down-regulating wound-suppressed transcripts (type IIa) or up-regulating wound-induced transcripts (type IIb). It is interesting that although all seven genes displayed their R-specific patterns in the treated tissues largely independently of the leaf or plant developmental stage, only the type I genes displayed strong systemic induction. Ethylene was not responsible for any of the specific patterns of expression. R collected from different tobacco feeding insects, M. sexta, Manduca quinquemaculata, and Heliothis virescens, as well as from different instars of M. sexta were equally active. The active components of M. sexta R were heat stable and active in minute amounts, comparable with real transfer rates during larval feeding. Specific expression patterns may indicate that the plant is adjusting its wound response to efficiently fend off M. sexta, but may also be advantageous to the larvae, especially when R suppress wound-induced plant responses.
Figures
References
-
- Alborn HT, Turlings TCJ, Jones TH, Stenhagen G, Loughrin JH, Tumlinson JH. An elicitor of plant volatiles from beet armyworm oral secretion. Science. 1997;276:945–949.
-
- Azevedo RA, Arruda P, Turner WL, Lea PJ. The biosynthesis and metabolism of the aspartate derived amino acids in higher plants. Phytochemistry. 1997;46:395–419. - PubMed
-
- Baker B, Zambryski P, Staskawicz B, Dinesh-Kumar SP. Signaling in plant-microbe interactions. Science. 1997;276:726–733. - PubMed
-
- Baldwin IT. The alkaloidal responses of wild tobacco to real and simulated herbivory. Oecologia. 1988;77:378–381. - PubMed
-
- Baldwin IT. Herbivory simulations in ecological research. Trends Ecol Evol. 1990;5:91–93. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
