

Variation between pathogenic serovars within Salmonella pathogenicity islands
- PMID: 12775700
- PMCID: PMC156220
- DOI: 10.1128/JB.185.12.3624-3635.2003
Variation between pathogenic serovars within Salmonella pathogenicity islands
Abstract
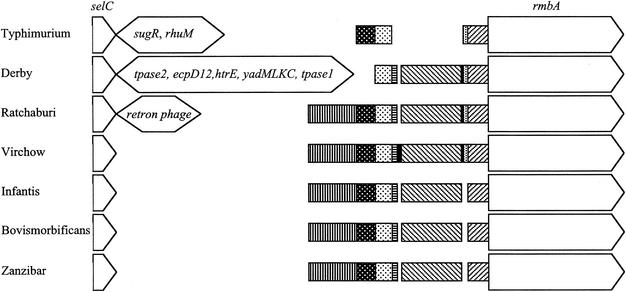
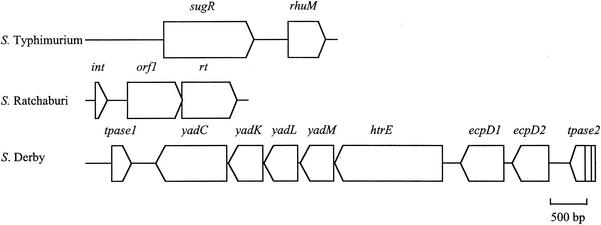
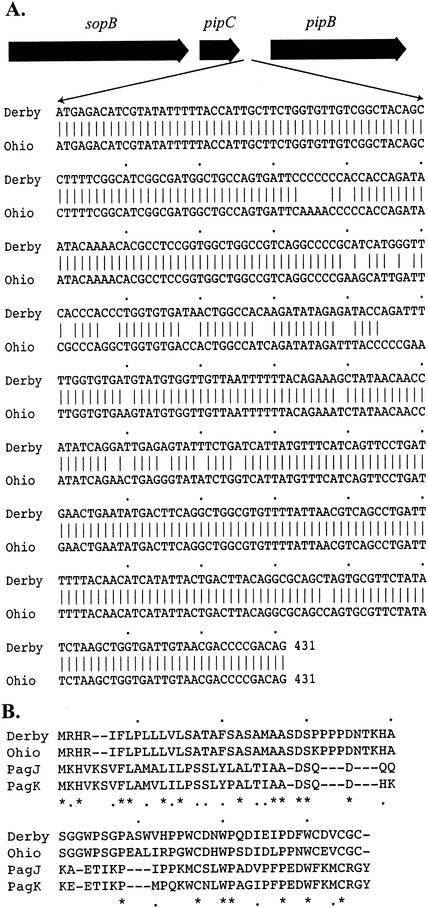
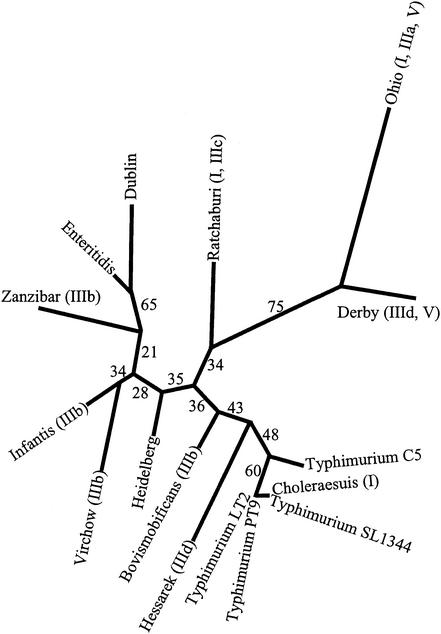
Although four of the five Salmonella pathogenicity islands (SPIs) have been characterized in detail for Salmonella enterica serovar Typhimurium, and the fifth has been characterized for Salmonella enterica serovar Dublin, there have been limited studies to examine them in detail in a range of pathogenic serovars of S. enterica. The aim of this study was to examine these regions, shown to be crucial in virulence, in pathogenic serovars to identify any major deletions or insertions that may explain variation in virulence and provide further understanding of the elements involved in the evolution of these regions. Multiple strains of each of the 13 serovars were compared by Southern blot hybridization using a series of probes that together encompassed the full length of all five SPIs. With the exception of serovar Typhimurium, all strains of the same serovar were identical in all five SPIs. Those serovars that differed from serovar Typhimurium in SPI-1 to SPI-4 and from serovar Dublin in SPI-5 were examined in more detail in the variant regions by PCR, and restriction endonuclease digestion and/or DNA sequencing. While most variation in hybridization patterns was attributable to loss or gain of single restriction endonuclease cleavage sites, three regions, in SPI-1, SPI-3, and SPI-5, had differences due to major insertions or deletions. In SPI-1 the avrA gene was replaced by a 200-base fragment in three serovars, as reported previously. In SPI-5, two serovars had acquired an insertion with similarity to the pagJ and pagK genes between pipC and pipD. In SPI-3 the genes sugR and rhuM were deleted in most serovars and in some were replaced by sequences that were very similar to either the Escherichia coli fimbrial operon, flanked by two distinct insertion sequence elements, or to the E. coli retron phage PhiR73. The distribution of these differences suggests that there have been a number of relatively recent horizontal transfers of genes into S. enterica and that in some cases the same event has occurred in multiple lineages of S. enterica. Thus, it seems that insertion sequences and retron phages are likely to be involved in continuing evolution of the pathogenicity islands of pathogenic Salmonella serovars.
Figures

References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Beltran, P., J. M. Musser, R. Helmuth, J. J. Farmer III, W. M. Frerichs, I. K. Wachsmuth, K. Ferris, A. C. McWhorter, J. G. Wells, A. Cravioto, et al. 1988. Toward a population genetic analysis of Salmonella: genetic diversity and relationships among strains of serotypes S. choleraesuis, S. derby, S. dublin, S. enteritidis, S. heidelberg, S. infantis, S. newport, and S. typhimurium. Proc. Natl. Acad. Sci. USA 85:7753-7757. - PMC - PubMed
-
- Boyd, E. F., F. S. Wang, P. Beltran, S. A. Plock, K. Nelson, and R. K. Selander. 1993. Salmonella reference collection B (SARB): strains of 37 serovars of subspecies I. J. Gen. Microbiol. 139:1125-1132. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
