

Cryptochromes define a novel circadian clock mechanism in monarch butterflies that may underlie sun compass navigation
- PMID: 18184036
- PMCID: PMC2174970
- DOI: 10.1371/journal.pbio.0060004
Cryptochromes define a novel circadian clock mechanism in monarch butterflies that may underlie sun compass navigation
Abstract
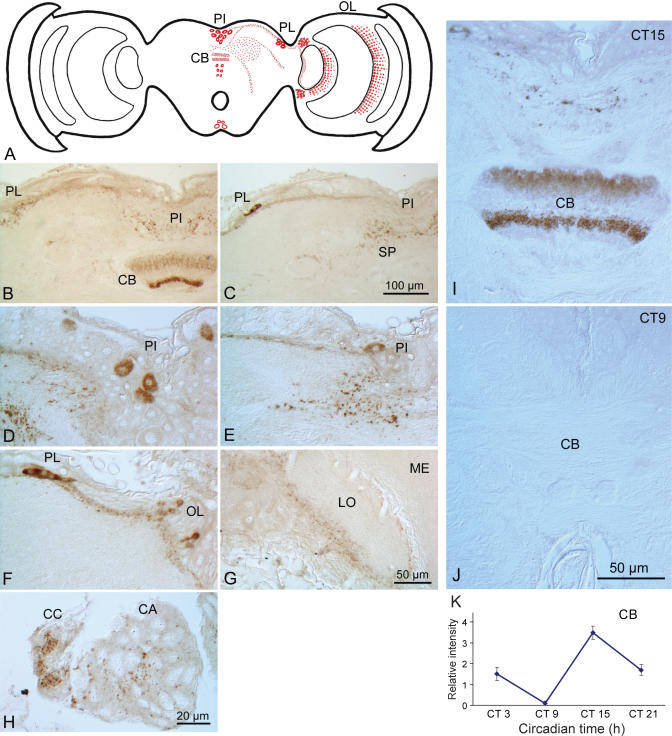
The circadian clock plays a vital role in monarch butterfly (Danaus plexippus) migration by providing the timing component of time-compensated sun compass orientation, a process that is important for successful navigation. We therefore evaluated the monarch clockwork by focusing on the functions of a Drosophila-like cryptochrome (cry), designated cry1, and a vertebrate-like cry, designated cry2, that are both expressed in the butterfly and by placing these genes in the context of other relevant clock genes in vivo. We found that similar temporal patterns of clock gene expression and protein levels occur in the heads, as occur in DpN1 cells, of a monarch cell line that contains a light-driven clock. CRY1 mediates TIMELESS degradation by light in DpN1 cells, and a light-induced TIMELESS decrease occurs in putative clock cells in the pars lateralis (PL) in the brain. Moreover, monarch cry1 transgenes partially rescue both biochemical and behavioral light-input defects in cry(b) mutant Drosophila. CRY2 is the major transcriptional repressor of CLOCK:CYCLE-mediated transcription in DpN1 cells, and endogenous CRY2 potently inhibits transcription without involvement of PERIOD. CRY2 is co-localized with clock proteins in the PL, and there it translocates to the nucleus at the appropriate time for transcriptional repression. We also discovered CRY2-positive neural projections that oscillate in the central complex. The results define a novel, CRY-centric clock mechanism in the monarch in which CRY1 likely functions as a blue-light photoreceptor for entrainment, whereas CRY2 functions within the clockwork as the transcriptional repressor of a negative transcriptional feedback loop. Our data further suggest that CRY2 may have a dual role in the monarch butterfly's brain-as a core clock element and as an output that regulates circadian activity in the central complex, the likely site of the sun compass.
Conflict of interest statement
Figures







Comment in
-
In monarchs, Cry2 is king of the clock.PLoS Biol. 2008 Jan;6(1):e12. doi: 10.1371/journal.pbio.0060012. Epub 2008 Jan 8. PLoS Biol. 2008. PMID: 20076689 Free PMC article. No abstract available.
References
-
- Saunders DS. Insect clocks. Boston: Elsevier; 2002. 560 xv,
-
- Stanewsky R. Genetic analysis of the circadian system in Drosophila melanogaster and mammals. J Neurobiol. 2003;54:111–147. - PubMed
-
- Williams JA, Sehgal A. Molecular components of the circadian system in Drosophila. Annu Rev Physiol. 2001;63:729–755. - PubMed
-
- Emery P, So WV, Kaneko M, Hall JC, Rosbash M. CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell. 1998;95:669–679. - PubMed
-
- Emery P, Stanewsky R, Helfrich-Forster C, Emery-Le M, Hall JC, et al. Drosophila CRY is a deep brain circadian photoreceptor. Neuron. 2000;26:493–504. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
