

Molecular marker systems for Oenothera genetics
- PMID: 18791241
- PMCID: PMC2581935
- DOI: 10.1534/genetics.108.091249
Molecular marker systems for Oenothera genetics
Abstract
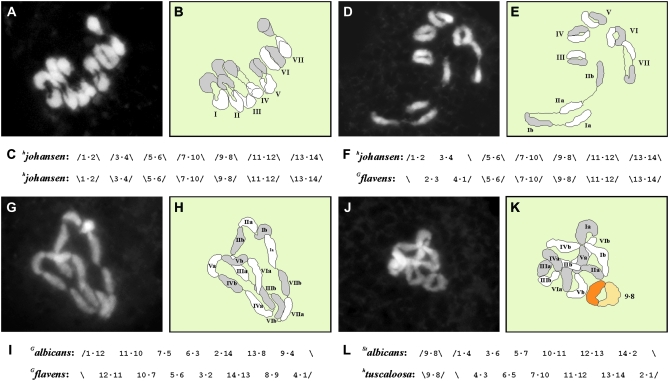
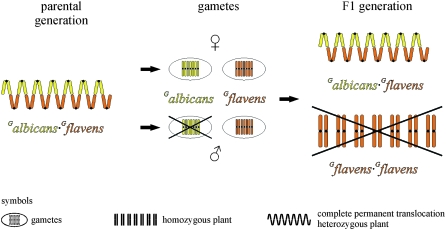
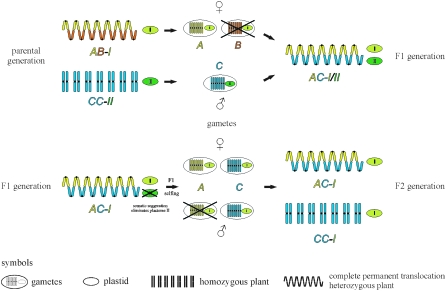
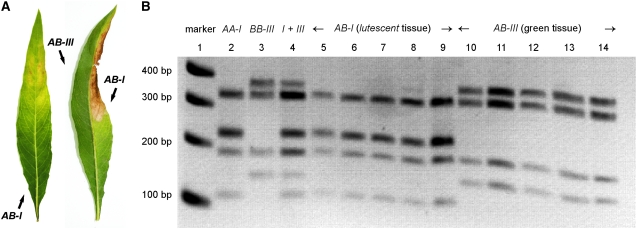
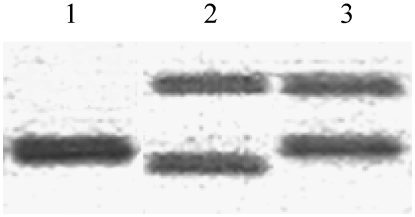
The genus Oenothera has an outstanding scientific tradition. It has been a model for studying aspects of chromosome evolution and speciation, including the impact of plastid nuclear co-evolution. A large collection of strains analyzed during a century of experimental work and unique genetic possibilities allow the exchange of genetically definable plastids, individual or multiple chromosomes, and/or entire haploid genomes (Renner complexes) between species. However, molecular genetic approaches for the genus are largely lacking. In this study, we describe the development of efficient PCR-based marker systems for both the nuclear genome and the plastome. They allow distinguishing individual chromosomes, Renner complexes, plastomes, and subplastomes. We demonstrate their application by monitoring interspecific exchanges of genomes, chromosome pairs, and/or plastids during crossing programs, e.g., to produce plastome-genome incompatible hybrids. Using an appropriate partial permanent translocation heterozygous hybrid, linkage group 7 of the molecular map could be assigned to chromosome 9.8 of the classical Oenothera map. Finally, we provide the first direct molecular evidence that homologous recombination and free segregation of chromosomes in permanent translocation heterozygous strains is suppressed.
Figures



References
-
- Barre, D. E., 2001. Potential of evening primrose, borage, black currant, and fungal oils in human health. Ann. Nutr. Metab. 45 47–57. - PubMed
-
- Birky, C. W., Jr., 2001. The inheritance of genes in mitochondria and chloroplasts: laws, mechanisms, and models. Annu. Rev. Genet. 35 125–148. - PubMed
-
- Burnham, C. R., 1962. Discussions in Cytogenetics. Burgess, Minneapolis.
-
- Chapman, M. J., D. L. Mulcahy and D. B. Stein, 1999. Identification of plastid genotypes in Oenothera subsect. Munzia by restriction fragment length polymorphisms (RFLPs). Theor. Appl. Genet. 98 47–53.
-
- Cleland, R. E., 1935. Cyto-taxonomic studies on certain Oenotheras from California. Proc. Am. Philos. Soc. 75 339–429.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
