

Genetic tools for allelic replacement in Burkholderia species
- PMID: 18502918
- PMCID: PMC2493169
- DOI: 10.1128/AEM.00531-08
Genetic tools for allelic replacement in Burkholderia species
Abstract
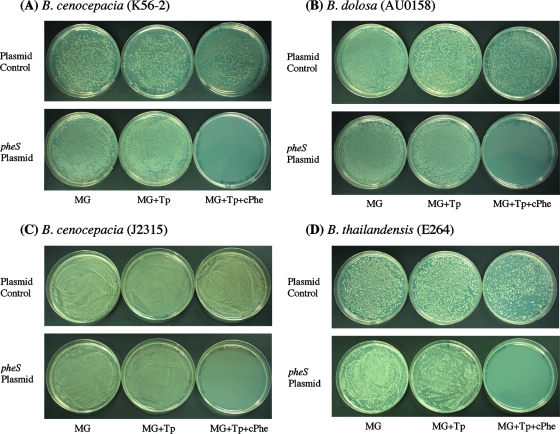
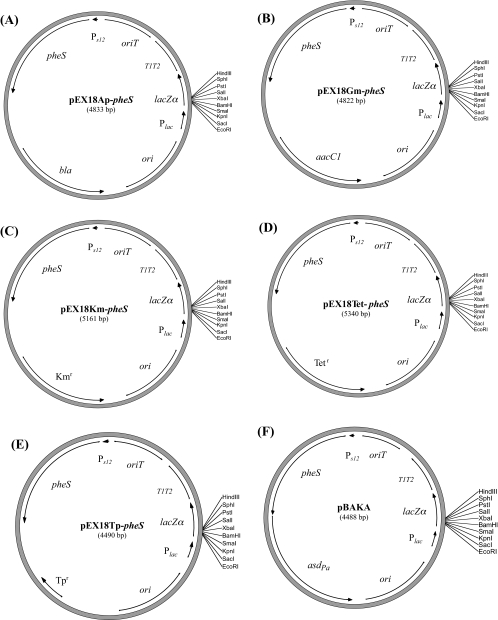
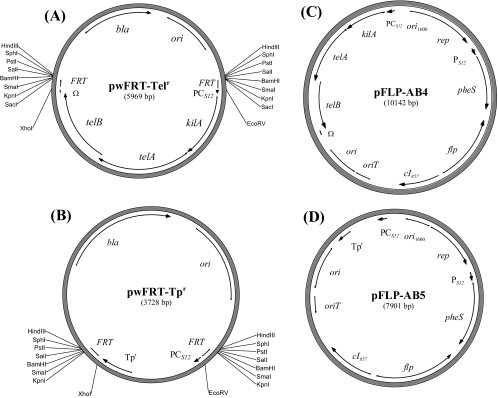
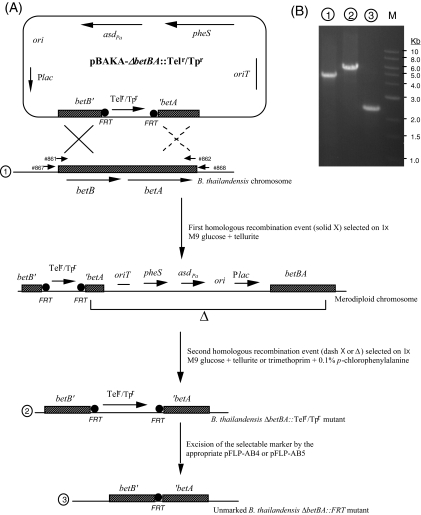
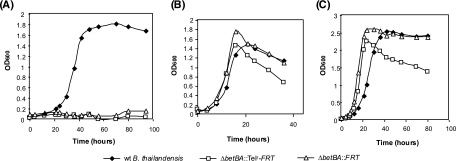
Allelic replacement in the Burkholderia genus has been problematic due to the lack of appropriate counter-selectable and selectable markers. The counter-selectable marker sacB, commonly used in gram-negative bacteria, is nonselective on sucrose in many Burkholderia species. In addition, the use of antibiotic resistance markers of clinical importance for the selection of desirable genetic traits is prohibited in the United States for two potential bioterrorism agents, Burkholderia mallei and Burkholderia pseudomallei. Here, we engineered a mutated counter-selectable marker based on the B. pseudomallei PheS (the alpha-subunit of phenylalanyl tRNA synthase) protein and tested its effectiveness in three different Burkholderia species. The mutant PheS protein effectively killed 100% of the bacteria in the presence of 0.1% p-chlorophenylalanine. We assembled the mutant pheS on several allelic replacement vectors, in addition to constructing selectable markers based on tellurite (Tel(r)) and trimethoprim (Tp(r)) resistance that are excisable by flanking unique FLP recombination target (FRT) sequences. As a proof of concept, we utilized one of these gene replacement vectors (pBAKA) and the Tel(r)-FRT cassette to produce a chromosomal mutation in the Burkholderia thailandensis betBA operon, which codes for betaine aldehyde dehydrogenase and choline dehydrogenase. Chromosomal resistance markers could be excised by the introduction of pFLP-AB5 (Tp(r)), which is one of two constructed flp-containing plasmids, pFLP-AB4 (Tel(r)) and pFLP-AB5 (Tp(r)). These flp-containing plasmids harbor the mutant pheS gene and allow self curing on media that contain p-chlorophenylalanine after Flp-FRT excision. The characterization of the Delta betBA::Tel(r)-FRT and Delta betBA::FRT mutants indicated a defect in growth with choline as a sole carbon source, while these mutants grew as well as the wild type with succinate and glucose as alternative carbon sources.
Figures
References
-
- Borgherini, G., P. Poubeau, F. Paganin, S. Picot, A. Michault, F. Thibault, and C. A. Berod. 2006. Melioidosis: an imported case from Madagascar. J. Travel Med. 13:318-320. - PubMed
-
- Brett, P. J., D. DeShazer, and D. E. Woods. 1998. Burkholderia thailandensis sp. nov., description of Burkholderia pseudomallei-like species. Int. J. Syst. Bacteriol. 48:317-320. - PubMed
-
- Brown, N. F., C.-A. Logue, J. A. Boddey, R. Scott, R. G. Hirst, and I. R. Beacham. 2004. Identification of a novel two-partner secretion system from Burkholderia pseudomallei. Mol. Gen. Genomics 272:204-215. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
