

Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees
- PMID: 19060168
- PMCID: PMC2632133
- DOI: 10.1128/AEM.02239-08
Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees
Abstract
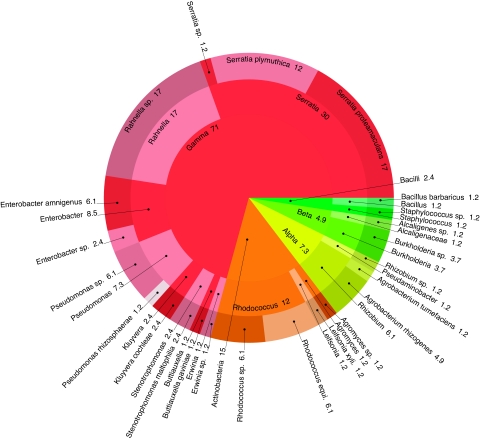
The association of endophytic bacteria with their plant hosts has a beneficial effect for many different plant species. Our goal is to identify endophytic bacteria that improve the biomass production and the carbon sequestration potential of poplar trees (Populus spp.) when grown in marginal soil and to gain an insight in the mechanisms underlying plant growth promotion. Members of the Gammaproteobacteria dominated a collection of 78 bacterial endophytes isolated from poplar and willow trees. As representatives for the dominant genera of endophytic gammaproteobacteria, we selected Enterobacter sp. strain 638, Stenotrophomonas maltophilia R551-3, Pseudomonas putida W619, and Serratia proteamaculans 568 for genome sequencing and analysis of their plant growth-promoting effects, including root development. Derivatives of these endophytes, labeled with gfp, were also used to study the colonization of their poplar hosts. In greenhouse studies, poplar cuttings (Populus deltoides x Populus nigra DN-34) inoculated with Enterobacter sp. strain 638 repeatedly showed the highest increase in biomass production compared to cuttings of noninoculated control plants. Sequence data combined with the analysis of their metabolic properties resulted in the identification of many putative mechanisms, including carbon source utilization, that help these endophytes to thrive within a plant environment and to potentially affect the growth and development of their plant hosts. Understanding the interactions between endophytic bacteria and their host plants should ultimately result in the design of strategies for improved poplar biomass production on marginal soils as a feedstock for biofuels.
Figures




References
-
- Arshad, M., and W. T. Frankenberger. 1991. Microbial production of plant hormones. Plant Soil. 133:1-8.
-
- Barac, T., S. Taghavi, B. Borremans, A. Provoost, L. Oeyen, J. V. Colpaert, J. Vangronsveld, and D. van der Lelie. 2004. Engineered endophytic bacteria improve phytoremediation of water-soluble, volatile, organic pollutants. Nat. Biotechnol. 22:583-588. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
