

doi: 10.1038/ng.245.
Epub 2008 Oct 26.
Mutations of LRTOMT, a fusion gene with alternative reading frames, cause nonsyndromic deafness in humans
Affiliations
- PMID: 18953341
- PMCID: PMC3404732
- DOI: 10.1038/ng.245
Item in Clipboard
Mutations of LRTOMT, a fusion gene with alternative reading frames, cause nonsyndromic deafness in humans
Nat Genet.
2008 Nov.
Abstract
Many proteins necessary for sound transduction have been identified through positional cloning of genes that cause deafness. We report here that mutations of LRTOMT are associated with profound nonsyndromic hearing loss at the DFNB63 locus on human chromosome 11q13.3-q13.4. LRTOMT has two alternative reading frames and encodes two different proteins, LRTOMT1 and LRTOMT2, detected by protein blot analyses. LRTOMT2 is a putative methyltransferase. During evolution, new transcripts can arise through partial or complete coalescence of genes. We provide evidence that in the primate lineage LRTOMT evolved from the fusion of two neighboring ancestral genes, which exist as separate genes (Lrrc51 and Tomt) in rodents.
Figures
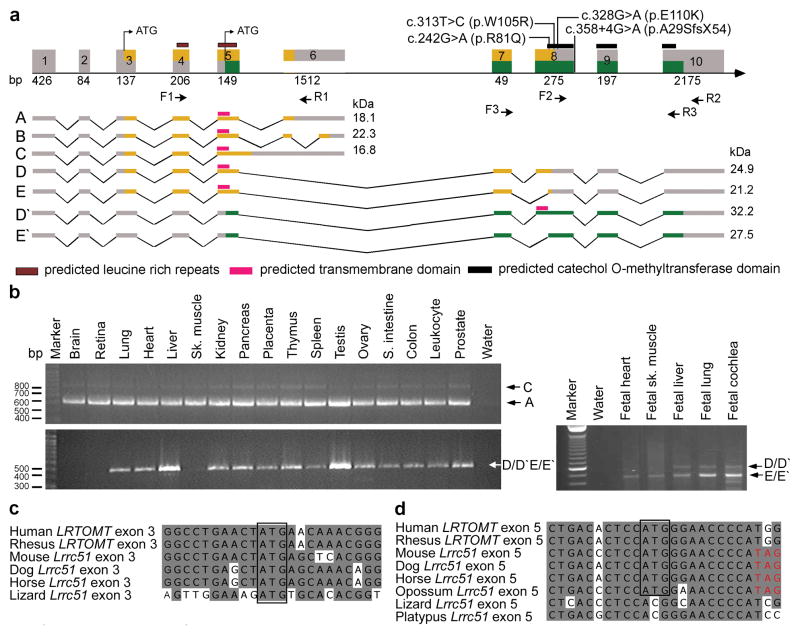
LRTOMT has alternative reading frames, and mutations cause nonsyndromic deafness. (a) LRTOMT has ten exons encoding multiple isoforms. Exons 5, 7 and 8 have dual reading frames. The two different reading frames of LRTOMT are colored orange and green. LRTOMT has two predicted translation start-codons, one in exon 3 and the second in exon 5. Grey boxes denote UTRs, and arrows show the location of primer-pairs for expression analyses. Isoforms A to E of LRTOMT1 have one predicted transmembrane domain (TM) and two leucine-rich repeats. Transcripts D′ and E′ are identical in sequence to D and E, respectively, but encode an entirely different protein, LRTOMT2, when translation starts in exon 5 and stops in exon 10. LRTOMT2 isoform D′ has a predicted catechol-O-methyltransferase domain and a TM. (b) PCR analyses of cDNAs from adult and fetal human tissues using primer-pairs shown in panel a. Transcripts A and C are amplified using primers F1 and R1 and are detected in all adult tissues tested. The transcripts D/D′ and E/E′ were detected either using RT-PCR primers F2 and R2 (adult tissues) or primers F3 and R3 (fetal tissues). (c-d) ClustalW alignments of nucleotide sequences of LRTOMT exons 3 and 5 and Lrrc51. Conserved translation start-codons of LRTOMT1 and LRTOMT2 are boxed. (d) If translation begins with the conserved ATG in exon 5 of mouse, dog, horse and opossum, there is an inframe translation stop-codon (TAG, red font).
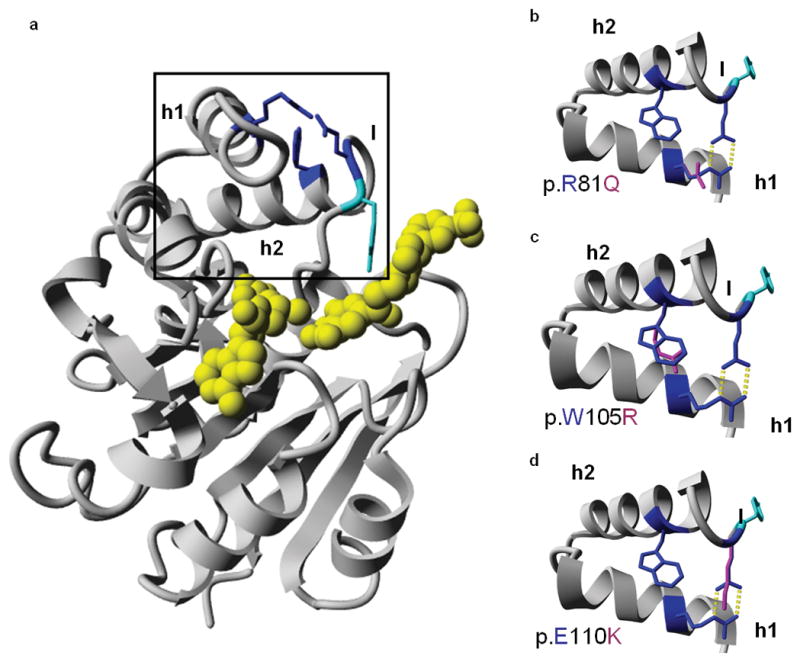
Molecular model and predicted effects of missense mutations. (a) Molecular model of the catechol-O-methyltransferase domain of LRTOMT2, residues 79-290. The mutated residues are depicted in blue. The predicted ligands are colored yellow, and the tyrosine residue (p.Y111) that lines the hydrophobic groove of the ligand binding site is shown in cyan. The region enlarged in b-d is boxed. (b-d) Missense mutations of LRTOMT2. The region of helices 1 and 2 and part of the flanking loops is enlarged. Wild type residues p.R81, p.W105 and p.E110 are depicted in blue, mutated residues in pink. Hydrogen bonds are represented by yellow dotted lines. (b) The p.R81 and p.E110 residues form a salt bridge between helix 1 and the loop following helix 2. The p.Q81 residue cannot form this salt bridge as it is not positively charged. Also, the formation of hydrogen bonds is impaired due to the smaller size of glutamine as compared to arginine. (c) The p.W105 residue is predicted to make hydrophobic interactions due to its big side chain. Most of these interactions would be lost by the p.W105R substitution. (d) Mutation p.E110K is predicted to lead to the loss of hydrogen bonds and a salt bridge. There would likely be repulsion between the side chains p.K110 and p.R81 since both are positively charged. h1, helix 1; h2, helix 2; l, loop
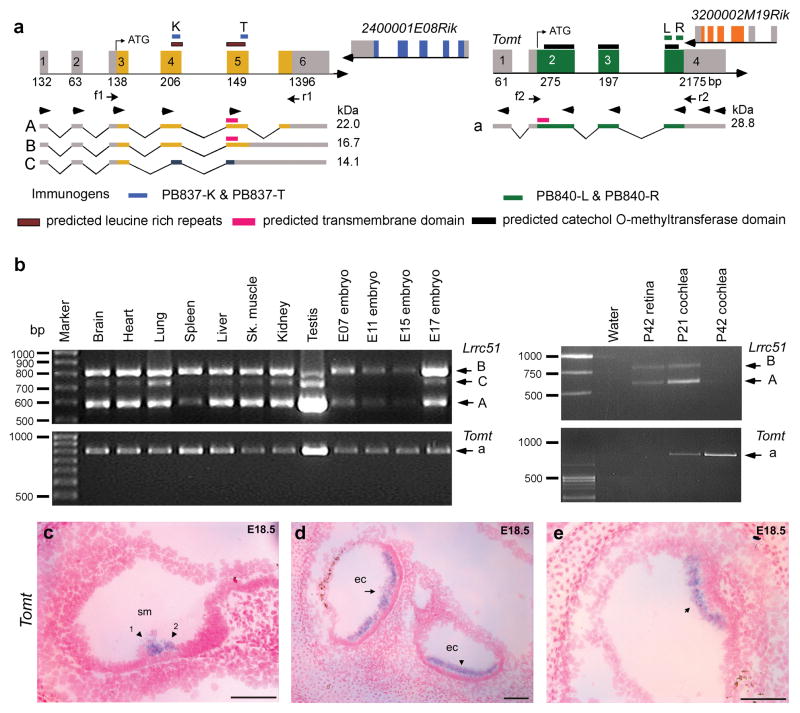
Mouse Lrrc51 and Tomt (a) Chromosomal region 7qE3 is syntenic to human chromosome 11q13.3. Unlike humans, mouse has two separate genes, Lrrc51 and Tomt encoding LRRC51 and TOMT, respectively. Translation of Tomt mRNA starts in exon 2. This ATG of LRTOMT is conserved in primates and located in human exon 8 (Supplementary Fig. 2d online). Right pointing arrowheads in exons 1–5 of Lrrc51 indicate forward RT-PCR primers used in all possible combinations with reverse primers (left pointing arrowheads) in Tomt and cDNAs from mouse brain, liver and heart. No mouse fusion transcripts were recovered (data not shown). Arrows (f1, r1 and f2, r2) indicate primer-pairs for expression profiling in b. (b) PCR analyses of Lrrc51 and Tomt transcripts show ubiquitous expression. (c-e) Tomt sense and antisense cRNA probes were hybridized to sagittal sections of whole mouse embryos from embryonic day 12.5 to 18.5. No signal was detected using the control sense probe (data not shown). (c) At E18.5 specific staining is visible in the region of the sensory cells of the cochlea where outer hair cells (arrowhead 1) and inner hair cells (arrowhead 2) are located. (d) At E18.5 in the utricle (arrow) and saccule (arrowhead), a clear signal can be observed in the region of the sensory cells. (e) In E18.5 sensory epithelium of the cristae ampullaris, Tomt mRNA was detected (arrow). No other tissues showed staining for Tomt at E18.5. Scale bars, 100 μm. sm, scala media; ec, endolymph compartment.
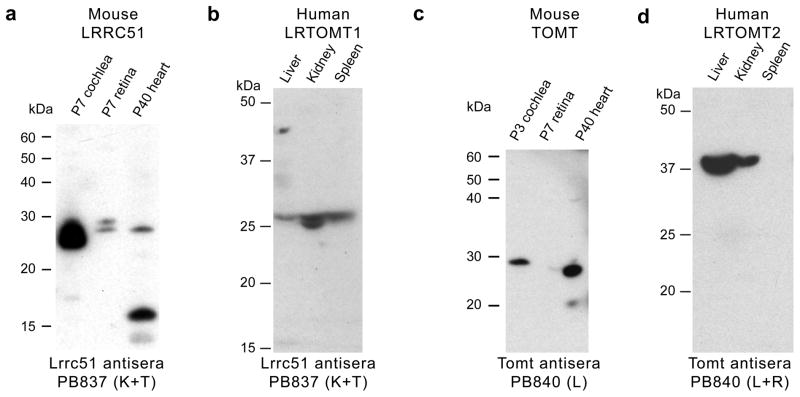
Western blot analyses of mouse LRRC51 and TOMT, and human LRTOMT1 and LRTOMT2. (a) Western blot analyses using anti-mouse LRRC51, PB837 (K+T) antisera and protein extracts from 7 day old (P7) mouse cochlea and retina (50 μg protein/lane) showed two bands of a size somewhat larger than the predicted sizes for LRRC51 protein isoforms A and B, while in P40 heart, the lower molecular weight isoform was detected along with a ~16 kDa band that might represent isoform C. (b) In human tissue, PB837 (K+T) detected proteins also of a size somewhat larger than predicted for human LRTOMT1 (isoform A, B and C). (c) Western blotting of anti-mouse TOMT (PB840-L) using affinity-purified antisera and mouse cochlear protein extracts (P3; 50 μg/lane) showed one band of about the expected size. In protein extracts from P40 heart, one ~28 kDa band of the expected deduced size (28.8 kDa) was detected. (d) Western blot analysis of protein from human tissues using anti-mouse TOMT antibodies (PB840 L+R) showed a signal in liver and kidney at ~37 kDa, slightly larger than the predicted size of 32.2 kDa for LRTOMT2 (isoform D′).
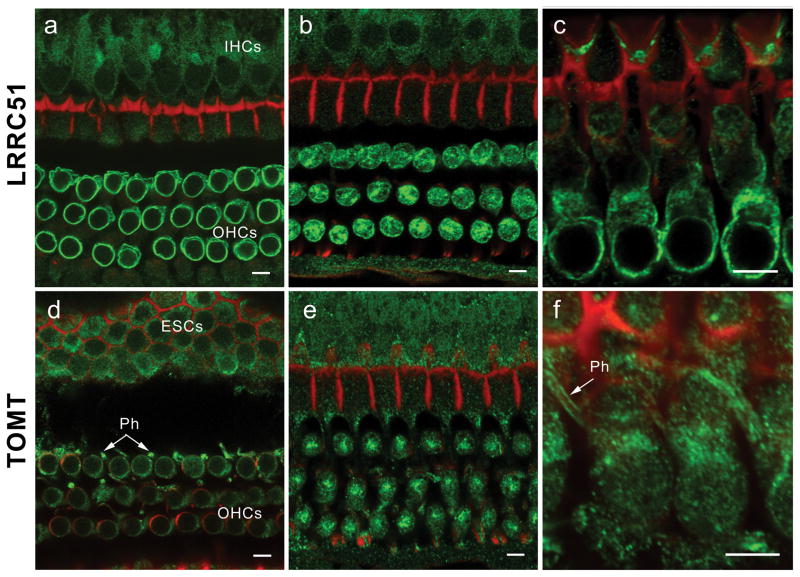
Immunolocalization of LRRC51 and TOMT in the P30 mouse inner ear (a) Anti-LRRC51 antiserum PB837 (green) immunostained the basolateral wall of the outer hair cells (OHCs), producing an annular fluorescence pattern in the optical cross-section of whole mount organ of Corti at the level of OHC nuclei. (b) Optical cross-section at the level below the cuticular plate of OHCs. Immunoreactivity to PB837-T antibody is observed at the lateral wall and in the cytoplasm of OHCs with a concentration at the site corresponding to the smooth endoplasmic reticulum. A weaker signal can also be observed in cytoplasm of inner hair cells (IHCs). (c) Longitudinal view of OHC bodies stained with PB837-K antibody highlighting the basolateral wall of OHCs. OHC nuclei are not stained. (d) Confocal images of the optical cross-section of the whole mount organ of Corti at the level of OHC nuclei immunostained with anti-mouse TOMT antibody. Cytoplasmic staining around the nuclei is seen, which is more evident in (e). Immunoreactivity is also observed in the cytoplasm of external sulcus cells (ESCs) and in phalanges (Ph) of outer phalangeal cells. (e) Optical cross-section at the level above the nuclei of OHCs. Immunoreactivity to PB840-L antibody in outer and inner hair cell bodies is concentrated under the cuticular plate of OHCs where smooth endoplasmic reticulum is located. (f) Longitudinal view of OHC bodies showing TOMT concentrated in the cytoplasm of OHCs above the nuclei. An arrow indicates staining of the phalanges (Ph) of outer phalangeal cells. The red signals represent rhodamine-phalloidin staining of F-actin. Scale bars, 5 μm.
Comment in
-
LRTOMT: a new tone in understanding the symphony of non-syndromic deafness.Clin Genet. 2009 Mar;75(3):227-9. doi: 10.1111/j.1399-0004.2009.01150_3.x. Clin Genet. 2009. PMID: 19250379 No abstract available.
References
-
- Friedman TB, Griffith AJ. Human nonsyndromic sensorineural deafness. Annu Rev Genomics Hum Genet. 2003;4:341–402. - PubMed
-
- Grant L, Fuchs PA. Auditory transduction in the mouse. Pflugers Arch. 2007;454:793–804. - PubMed
-
- Morton CC, Nance WE. Newborn hearing screening--a silent revolution. N Engl J Med. 2006;354:2151–64. - PubMed
-
- Pasek S, Risler JL, Brezellec P. Gene fusion/fission is a major contributor to evolution of multi-domain bacterial proteins. Bioinformatics. 2006;22:1418–23. - PubMed
-
- Kalay E, et al. A novel locus for autosomal recessive nonsyndromic hearing impairment, DFNB63, maps to chromosome 11q13.2–q13.4. J Mol Med. 2007;85:397–404. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
