

Secreted transcription factor controls Mycobacterium tuberculosis virulence
- PMID: 18685700
- PMCID: PMC2862998
- DOI: 10.1038/nature07219
Secreted transcription factor controls Mycobacterium tuberculosis virulence
Abstract
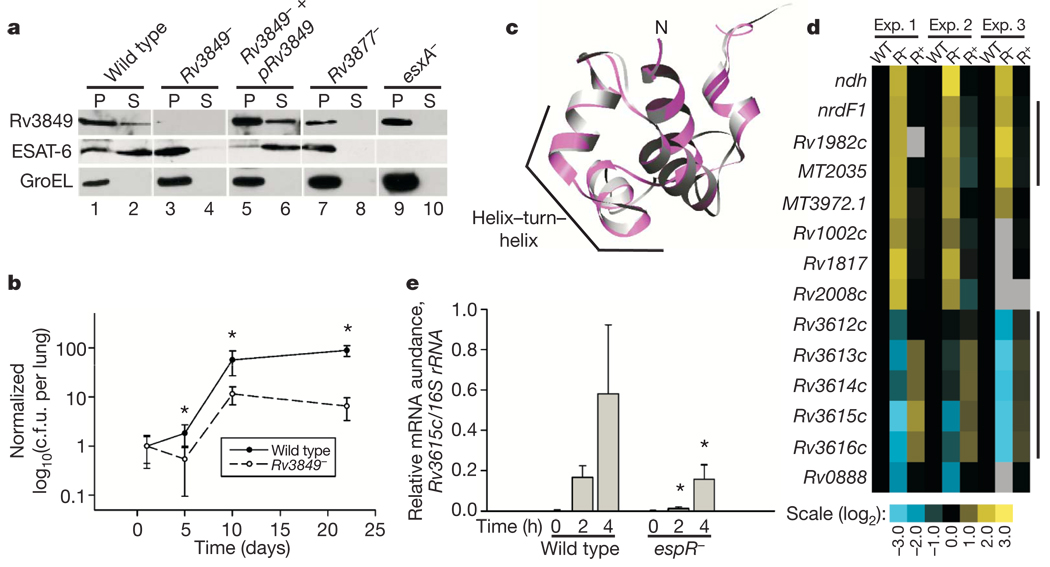
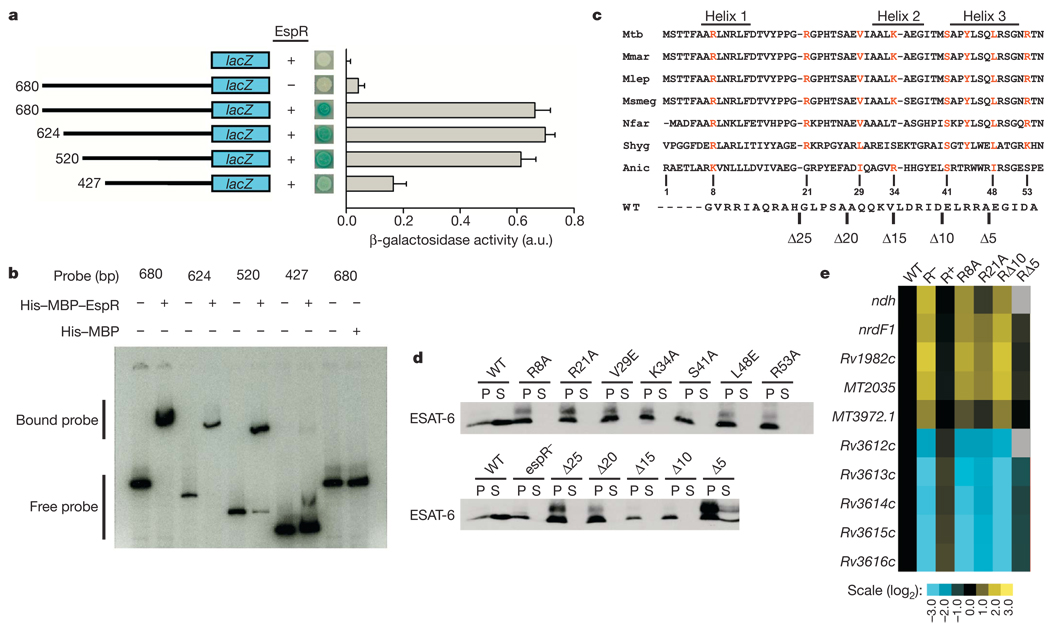
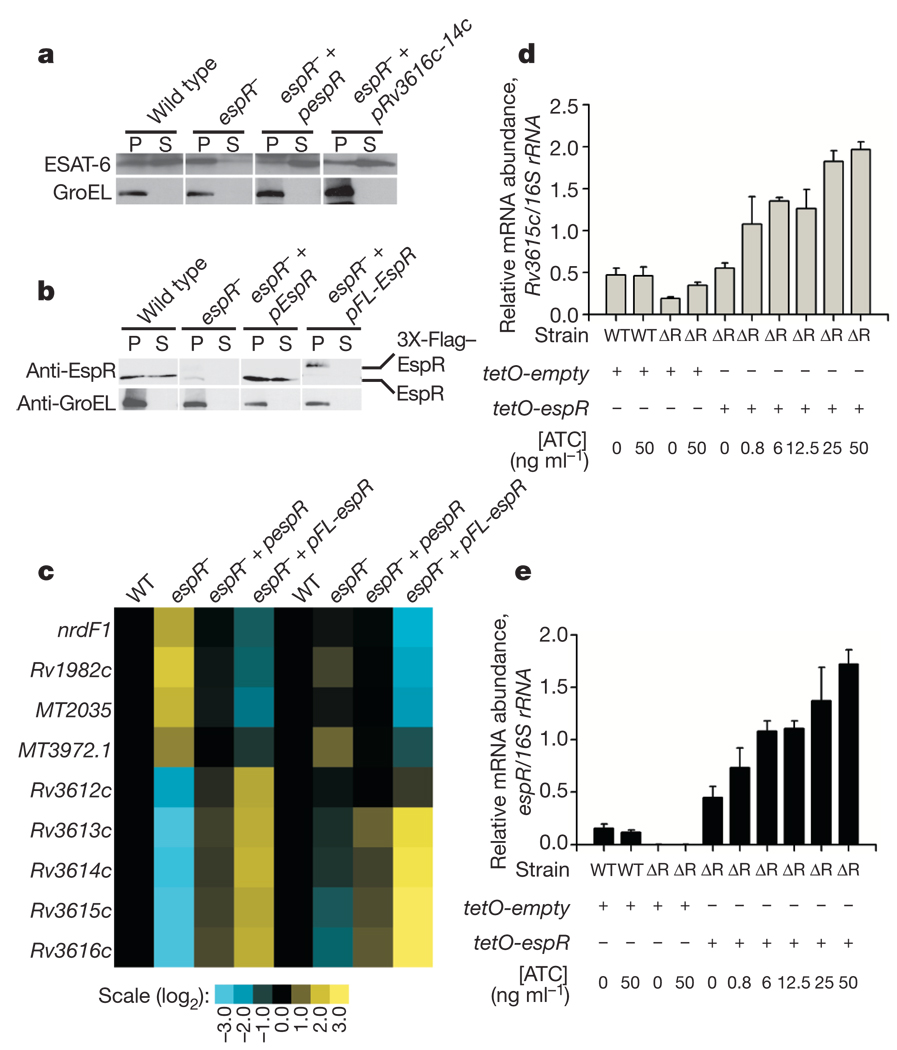
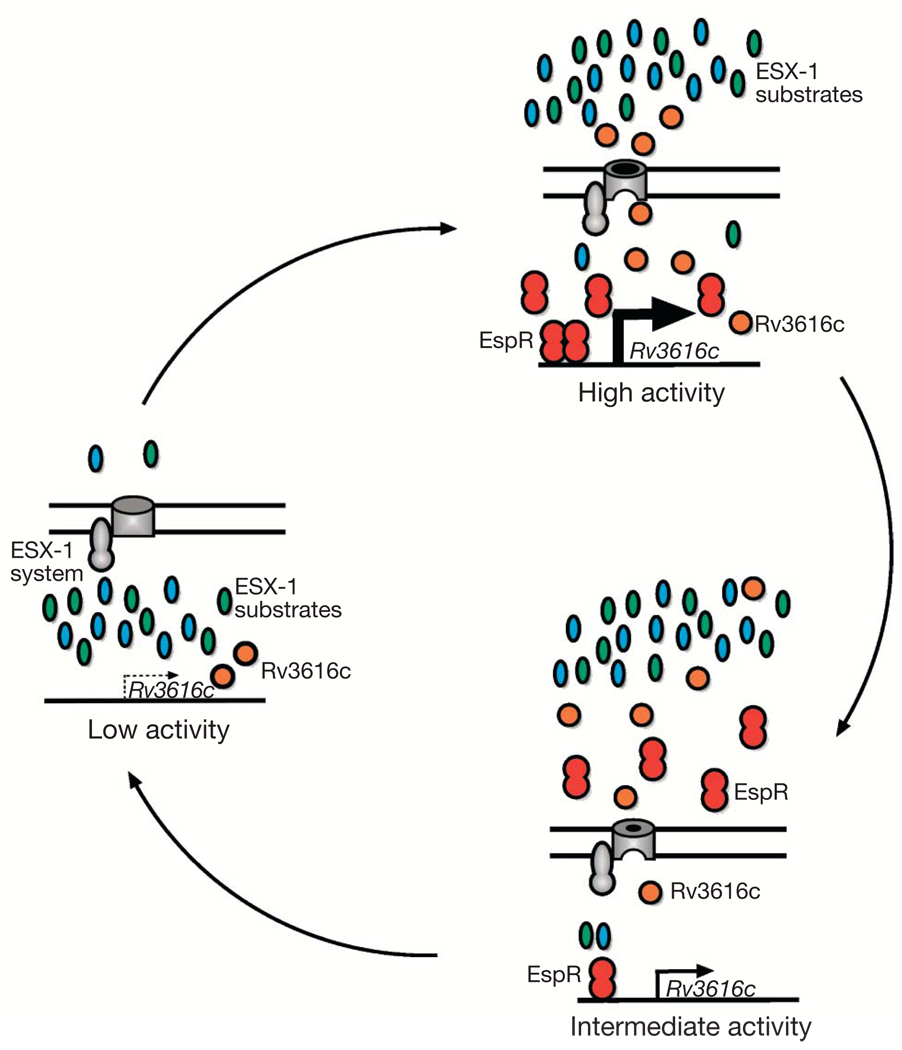
Bacterial pathogens trigger specialized virulence factor secretion systems on encountering host cells. The ESX-1 protein secretion system of Mycobacterium tuberculosis-the causative agent of the human disease tuberculosis-delivers bacterial proteins into host cells during infection and is critical for virulence, but how it is regulated is unknown. Here we show that EspR (also known as Rv3849) is a key regulator of ESX-1 that is required for secretion and virulence in mice. EspR activates transcription of an operon that includes three ESX-1 components, Rv3616c-Rv3614c, whose expression in turn promotes secretion of ESX-1 substrates. EspR directly binds to and activates the Rv3616c-Rv3614c promoter and, unexpectedly, is itself secreted from the bacterial cell by the ESX-1 system that it regulates. Efflux of the DNA-binding regulator results in reduced Rv3616c-Rv3614c transcription, and thus reduced ESX-1 secretion. Our results reveal a direct negative feedback loop that regulates the activity of a secretion system essential for virulence. As the virulence factors secreted by the ESX-1 system are highly antigenic, fine control of secretion may be critical to successful infection.
Figures
Comment in
-
Tuberculosis: Shrewd survival strategy.Nature. 2008 Aug 7;454(7205):702-3. doi: 10.1038/454702a. Nature. 2008. PMID: 18685690 No abstract available.
References
-
- Pathak SK, et al. Direct extracellular interaction between the early secreted antigen ESAT-6 of Mycobacterium tuberculosis and TLR2 inhibits TLR signaling in macrophages. Nature Immunol. 2007;8:610–618. - PubMed
-
- Abdallah AM, et al. Type VII secretion-mycobacteria show the way. Nature Rev. Microbiol. 2007;5:883–891. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
