

A protracted and dynamic maturation schedule underlies Arabidopsis leaf development
- PMID: 18805992
- PMCID: PMC2570723
- DOI: 10.1105/tpc.107.057521
A protracted and dynamic maturation schedule underlies Arabidopsis leaf development
Abstract
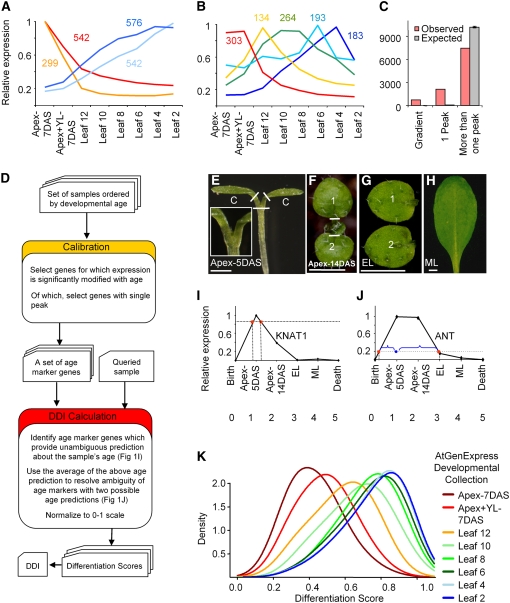
Leaf development has been monitored chiefly by following anatomical markers. Analysis of transcriptome dynamics during leaf maturation revealed multiple expression patterns that rise or fall with age or that display age-specific peaks. These were used to formulate a digital differentiation index (DDI) based on a set of selected markers with informative expression during leaf ontogeny. The leaf-based DDI reliably predicted the developmental state of leaf samples from diverse sources and was independent of mitotic cell division transcripts or propensity of specific cell types. When calibrated by informative root markers, the same algorithm accurately diagnosed dissected root samples. We used the DDI to characterize plants with reduced activities of multiple CINCINNATA (CIN)-TCP (TEOSINTE BRANCHED1, CYCLOIDEA, PCF) growth regulators. These plants had giant curled leaves made up of small cells with abnormal shape, low DDI scores, and low expression of mitosis markers, depicting the primary role of CIN-TCPs as promoters of differentiation. Delayed activity of several CIN-TCPs resulted in abnormally large but flat leaves with regular cells. The application of DDI has therefore portrayed the CIN-TCPs as heterochronic regulators that permit the development of a flexible and robust leaf form through an ordered and protracted maturation schedule.
Figures




References
-
- Alonso, J.M., and Stepanove, A.N. (2003). T-DNA mutagenesis in Arabidopsis. Methods Mol. Biol. 236 177–188. - PubMed
-
- Avery, G.S. (1933). Structure and development of the tobacco leaf. Am. J. Bot. 20 565–592.
-
- Birnbaum, K., Shasha, D.E., Wang, J.Y., Jung, J.W., Lambert, G.M., Galbraith, D.W., and Benfey, P.N. (2003). A gene expression map of the Arabidopsis root. Science 302 1956–1960. - PubMed
-
- Brady, S.M., Orlando, D.A., Lee, J.Y., Wang, J.Y., Koch, J., Dinneny, J.R., Mace, D., Ohler, U., and Benfey, P.N. (2007). A high-resolution root spatiotemporal map reveals dominant expression patterns. Science 318 801–806. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
