

Computational identification of gene-social environment interaction at the human IL6 locus
- PMID: 20176930
- PMCID: PMC2851818
- DOI: 10.1073/pnas.0911515107
Computational identification of gene-social environment interaction at the human IL6 locus
Abstract
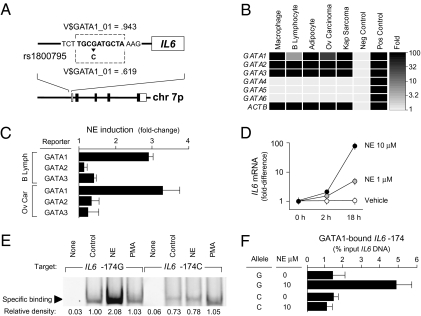
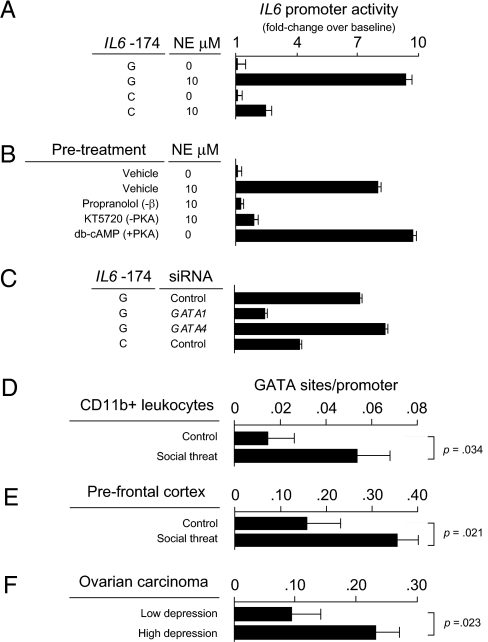
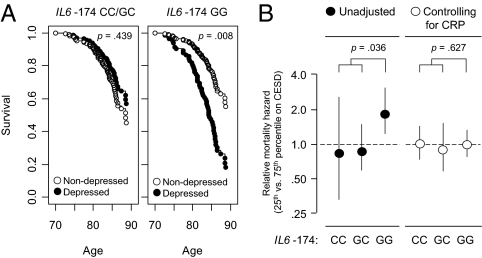
To identify genetic factors that interact with social environments to impact human health, we used a bioinformatic strategy that couples expression array-based detection of environmentally responsive transcription factors with in silico discovery of regulatory polymorphisms to predict genetic loci that modulate transcriptional responses to stressful environments. Tests of one predicted interaction locus in the human IL6 promoter (SNP rs1800795) verified that it modulates transcriptional response to beta-adrenergic activation of the GATA1 transcription factor in vitro. In vivo validation studies confirmed links between adverse social conditions and increased transcription of GATA1 target genes in primary neural, immune, and cancer cells. Epidemiologic analyses verified the health significance of those molecular interactions by documenting increased 10-year mortality risk associated with late-life depressive symptoms that occurred solely for homozygous carriers of the GATA1-sensitive G allele of rs1800795. Gating of depression-related mortality risk by IL6 genotype pertained only to inflammation-related causes of death and was associated with increased chronic inflammation as indexed by plasma C-reactive protein. Computational modeling of molecular interactions, in vitro biochemical analyses, in vivo animal modeling, and human molecular epidemiologic analyses thus converge in identifying beta-adrenergic activation of GATA1 as a molecular pathway by which social adversity can alter human health risk selectively depending on individual genetic status at the IL6 locus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Robinson GE. Genomics. Beyond nature and nurture. Science. 2004;304:397–399. - PubMed
-
- Hernandez LM, Blazer DG. Washington, DC: National Academies Press; 2006. - PubMed
-
- Caspi A, Moffitt TE. Gene-environment interactions in psychiatry: Joining forces with neuroscience. Nat Rev Neurosci. 2006;7:583–590. - PubMed
-
- Finch CE. The Biology of Human Longevity: Inflammation, Nutrition, and Aging in the Evolution of Lifespans. Burlington, MA: Academic Press; 2007.
-
- Meaney MJ, Szyf M, Seckl JR. Epigenetic mechanisms of perinatal programming of hypothalamic-pituitary-adrenal function and health. Trends Mol Med. 2007;13:269–277. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
- R01 AI052737/AI/NIAID NIH HHS/United States
- R01 CA109298/CA/NCI NIH HHS/United States
- R01 MH046801/MH/NIMH NIH HHS/United States
- CA110793/CA/NCI NIH HHS/United States
- AG107265/AG/NIA NIH HHS/United States
- R01 CA104825/CA/NCI NIH HHS/United States
- P30 AG028748/AG/NIA NIH HHS/United States
- AI052737/AI/NIAID NIH HHS/United States
- MH046801/MH/NIMH NIH HHS/United States
- CA116778/CA/NCI NIH HHS/United States
- CA109298/CA/NCI NIH HHS/United States
- R01 CA116778/CA/NCI NIH HHS/United States
- P60 AG010415/AG/NIA NIH HHS/United States
- R01 CA110793/CA/NCI NIH HHS/United States
- AG010415/AG/NIA NIH HHS/United States
- AG028748/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
