

Protection from obesity and diabetes by blockade of TGF-β/Smad3 signaling
- PMID: 21723505
- PMCID: PMC3169298
- DOI: 10.1016/j.cmet.2011.04.013
Protection from obesity and diabetes by blockade of TGF-β/Smad3 signaling
Abstract
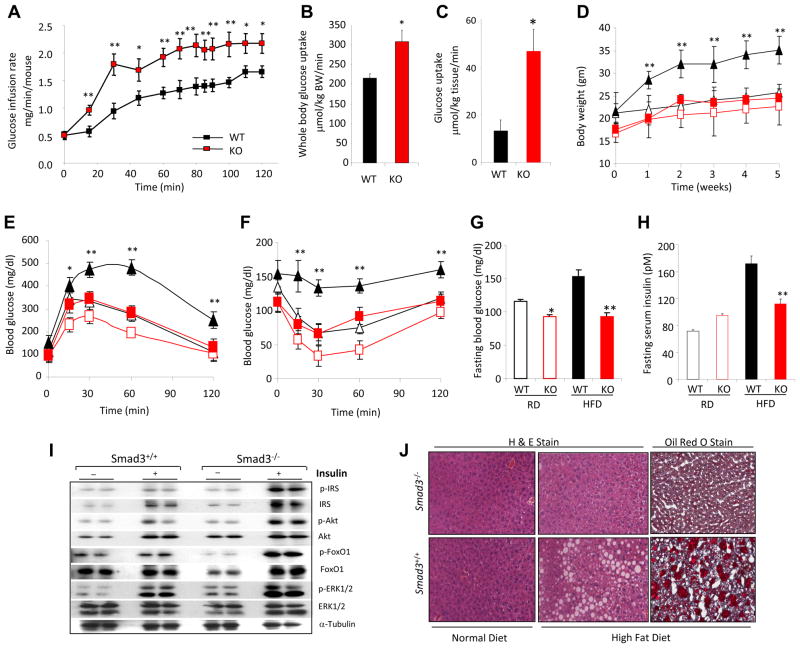
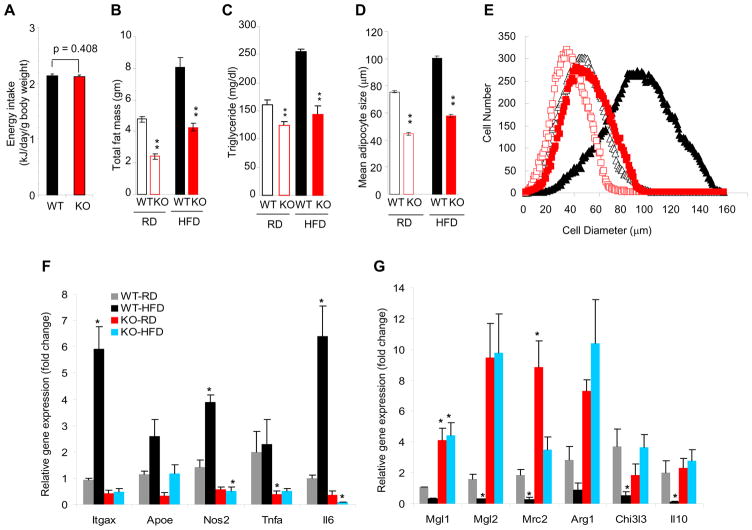
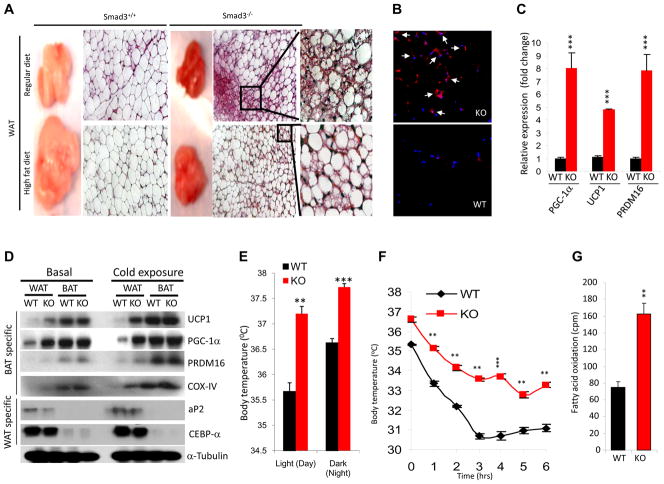
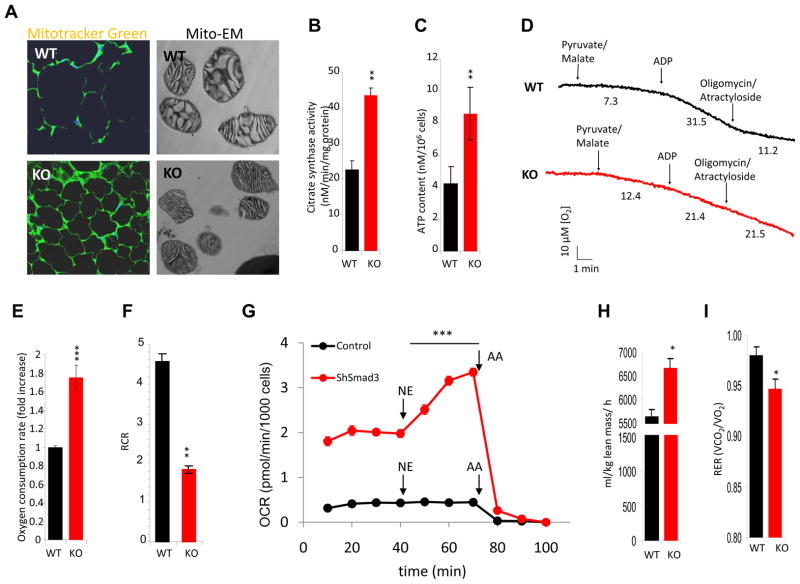
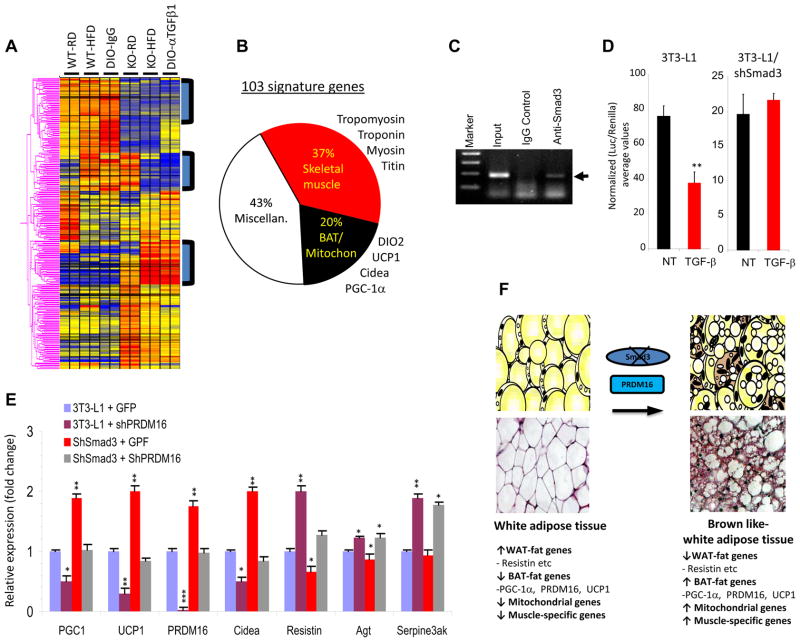
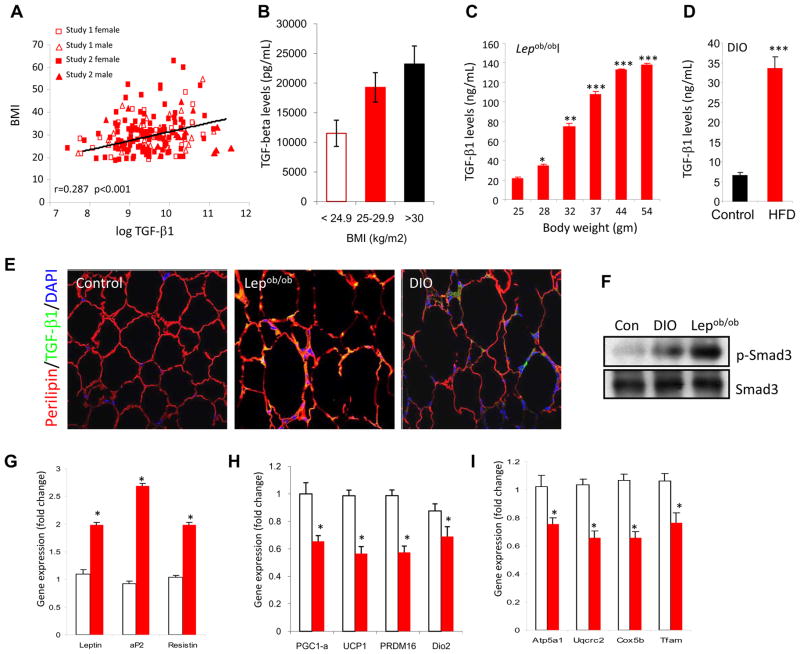
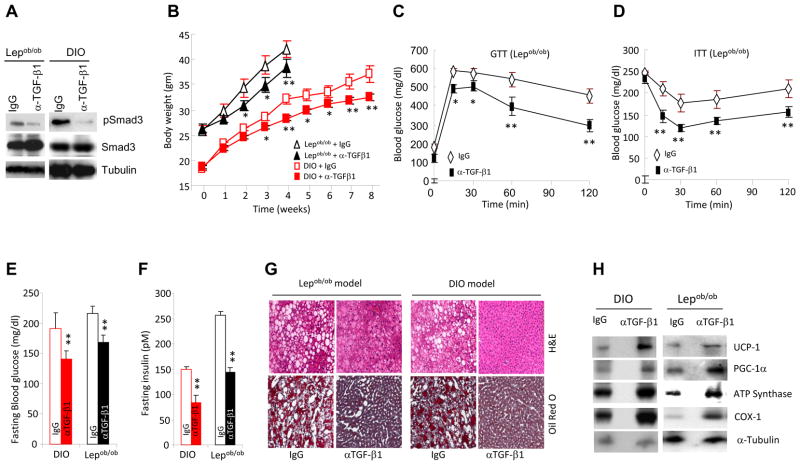
Imbalances in glucose and energy homeostasis are at the core of the worldwide epidemic of obesity and diabetes. Here, we illustrate an important role of the TGF-β/Smad3 signaling pathway in regulating glucose and energy homeostasis. Smad3-deficient mice are protected from diet-induced obesity and diabetes. Interestingly, the metabolic protection is accompanied by Smad3(-)(/-) white adipose tissue acquiring the bioenergetic and gene expression profile of brown fat/skeletal muscle. Smad3(-/-) adipocytes demonstrate a marked increase in mitochondrial biogenesis, with a corresponding increase in basal respiration, and Smad3 acts as a repressor of PGC-1α expression. We observe significant correlation between TGF-β1 levels and adiposity in rodents and humans. Further, systemic blockade of TGF-β signaling protects mice from obesity, diabetes, and hepatic steatosis. Together, these results demonstrate that TGF-β signaling regulates glucose tolerance and energy homeostasis and suggest that modulation of TGF-β activity might be an effective treatment strategy for obesity and diabetes.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Metabolic disease: Turning 'bad' fat into 'good'.Nat Rev Drug Discov. 2011 Aug 31;10(9):659. doi: 10.1038/nrd3540. Nat Rev Drug Discov. 2011. PMID: 21878978 No abstract available.
References
-
- Alessi MC, Bastelica D, Morange P, Berthet B, Leduc I, Verdier M, Geel O, Juhan-Vague I. Plasminogen activator inhibitor 1, transforming growth factor-beta1, and BMI are closely associated in human adipose tissue during morbid obesity. Diabetes. 2000;49:1374–1380. - PubMed
-
- Atit R, Sgaier SK, Mohamed OA, Taketo MM, Dufort D, Joyner AL, Niswander L, Conlon RA. Beta-catenin activation is necessary and sufficient to specify the dorsal dermal fate in the mouse. Developmental biology. 2006;296:164–176. - PubMed
-
- Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiological reviews. 2004;84:277–359. - PubMed
-
- Cannon B, Nedergaard J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. The Journal of experimental biology. 2011;214:242–253. - PubMed
-
- Choy L, Derynck R. Transforming growth factor-beta inhibits adipocyte differentiation by Smad3 interacting with CCAAT/enhancer-binding protein (C/EBP) and repressing C/EBP transactivation function. The Journal of biological chemistry. 2003;278:9609–9619. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
