

Lactate racemase is a nickel-dependent enzyme activated by a widespread maturation system
- PMID: 24710389
- PMCID: PMC4066177
- DOI: 10.1038/ncomms4615
Lactate racemase is a nickel-dependent enzyme activated by a widespread maturation system
Abstract
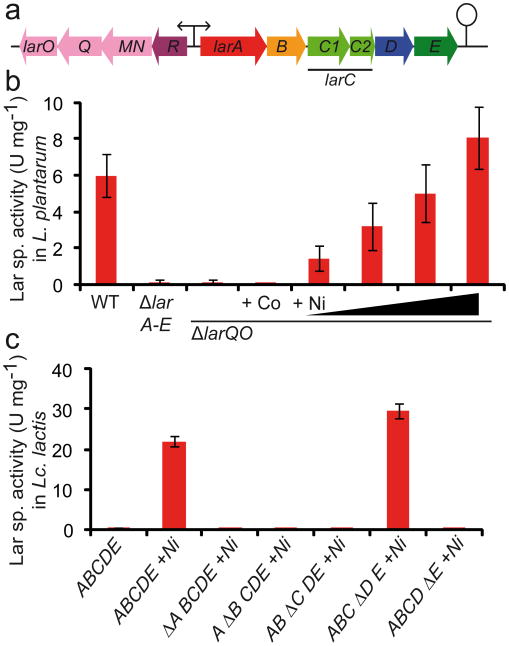
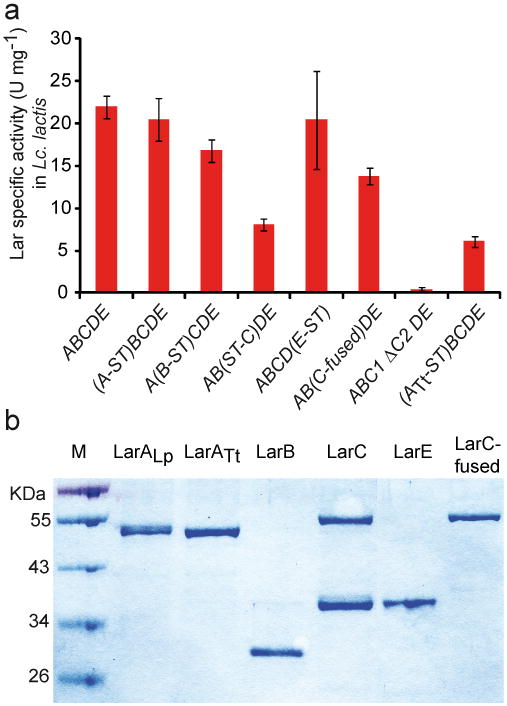
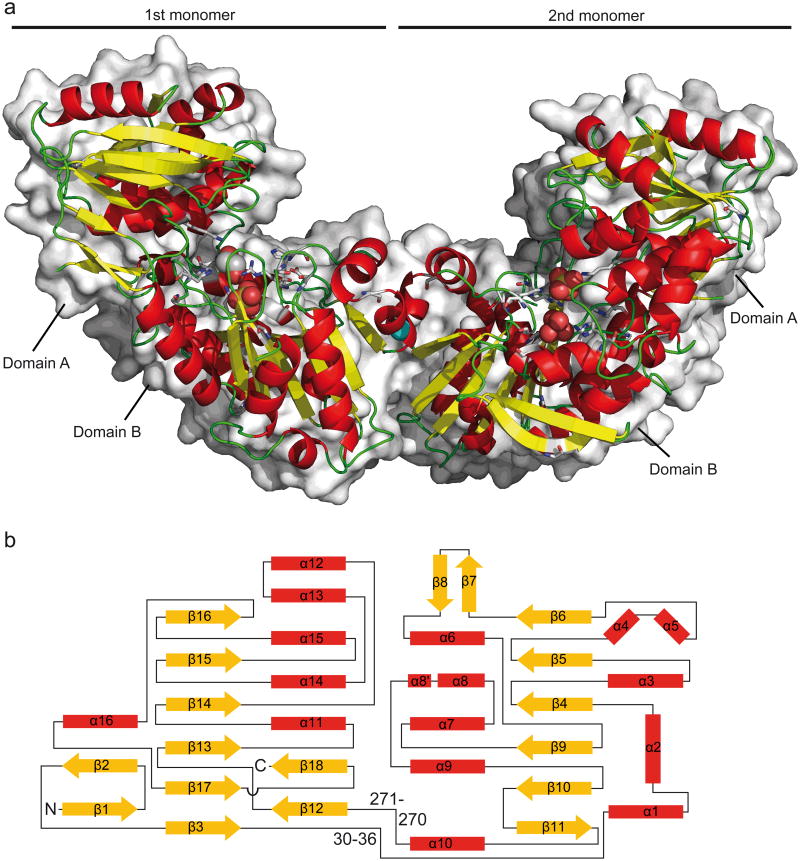
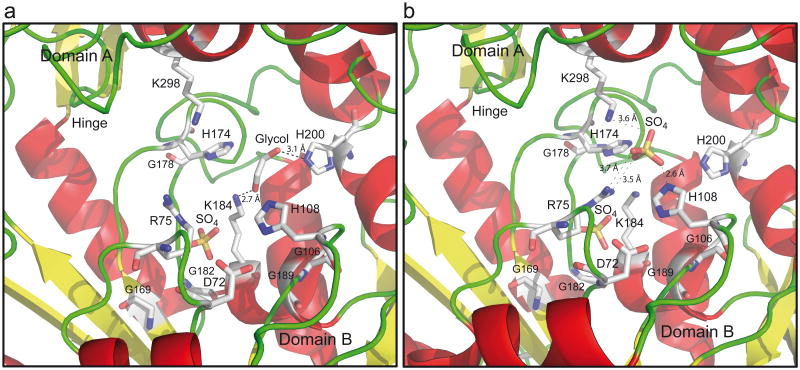
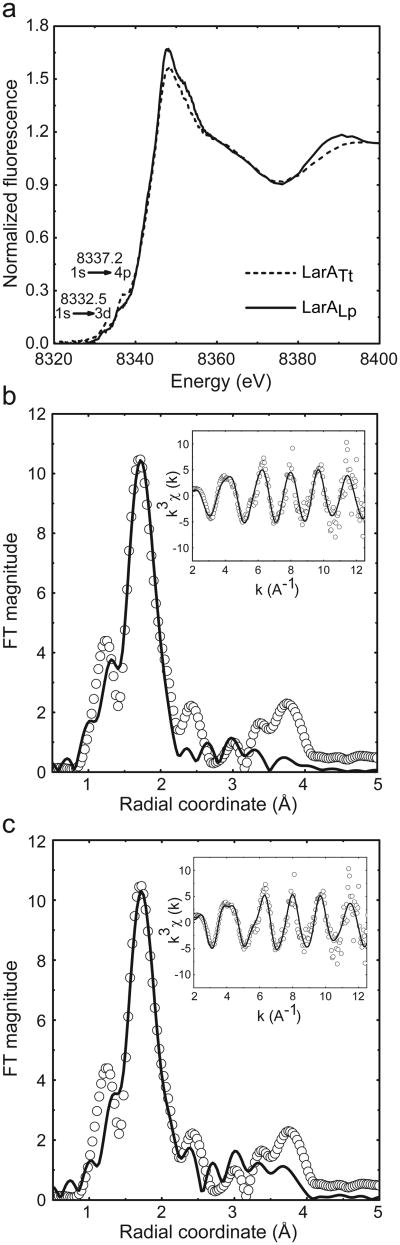
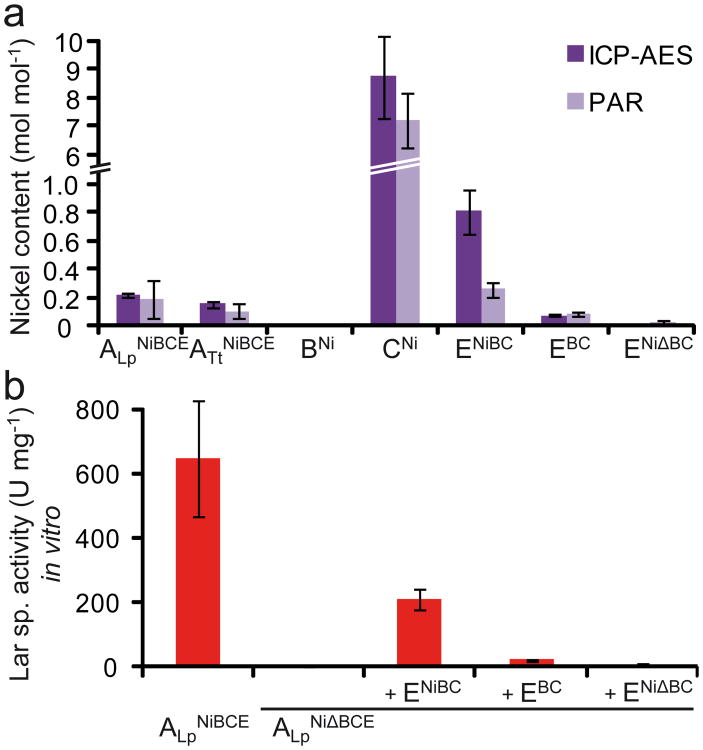
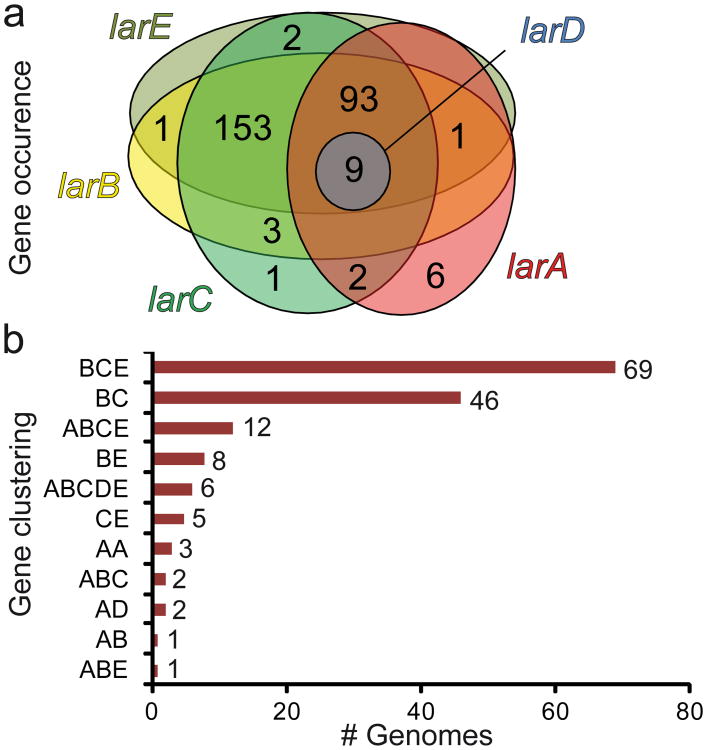
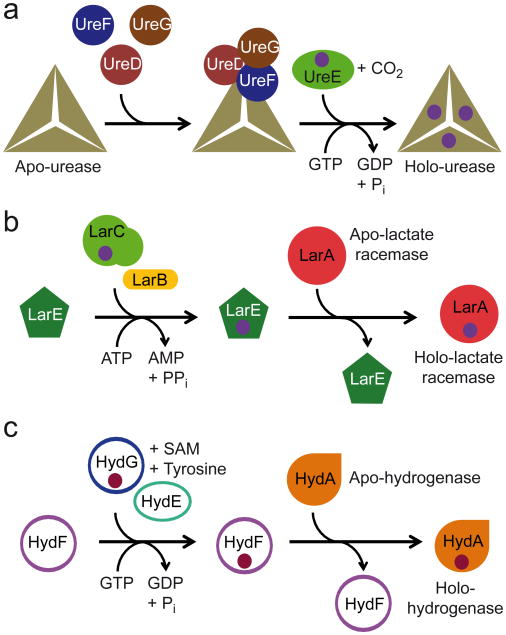
Racemases catalyse the inversion of stereochemistry in biological molecules, giving the organism the ability to use both isomers. Among them, lactate racemase remains unexplored due to its intrinsic instability and lack of molecular characterization. Here we determine the genetic basis of lactate racemization in Lactobacillus plantarum. We show that, unexpectedly, the racemase is a nickel-dependent enzyme with a novel α/β fold. In addition, we decipher the process leading to an active enzyme, which involves the activation of the apo-enzyme by a single nickel-containing maturation protein that requires preactivation by two other accessory proteins. Genomic investigations reveal the wide distribution of the lactate racemase system among prokaryotes, showing the high significance of both lactate enantiomers in carbon metabolism. The even broader distribution of the nickel-based maturation system suggests a function beyond activation of the lactate racemase and possibly linked with other undiscovered nickel-dependent enzymes.
Figures
References
-
- Okano K, Tanaka T, Ogino C, Fukuda H, Kondo A. Biotechnological production of enantiomeric pure lactic acid from renewable resources: recent achievements, perspectives, and limits. Appl Microbiol Biotechnol. 2010;85:413–423. - PubMed
-
- Walsh CT. Vancomycin resistance: decoding the molecular logic. Science. 1993;261:308–309. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
