

Efficient utilization of complex N-linked glycans is a selective advantage for Bacteroides fragilis in extraintestinal infections
- PMID: 25139987
- PMCID: PMC4156730
- DOI: 10.1073/pnas.1407344111
Efficient utilization of complex N-linked glycans is a selective advantage for Bacteroides fragilis in extraintestinal infections
Abstract
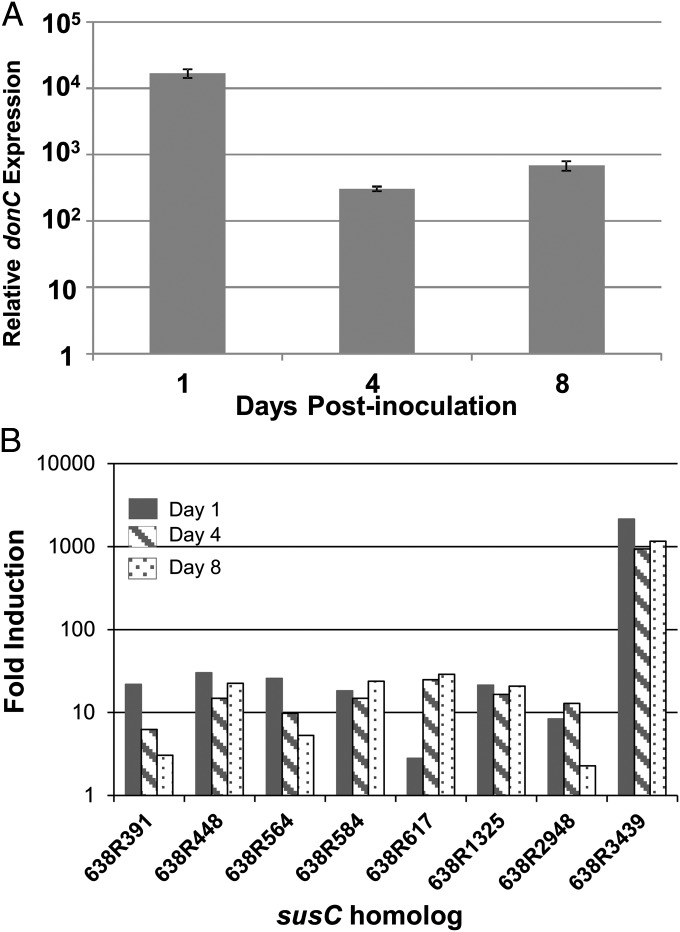
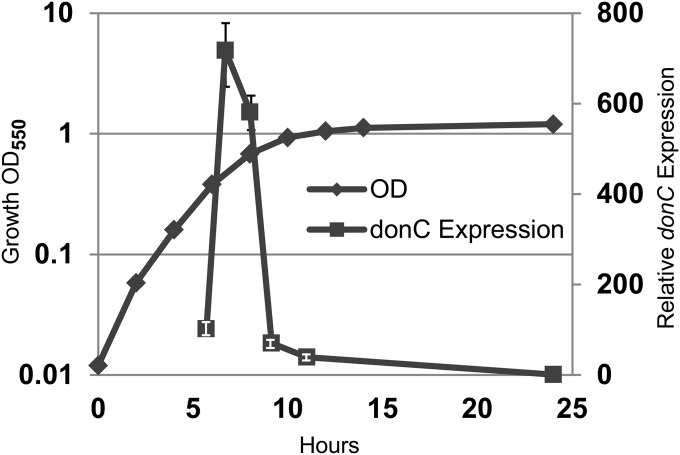
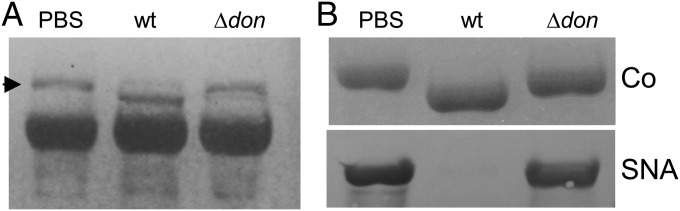
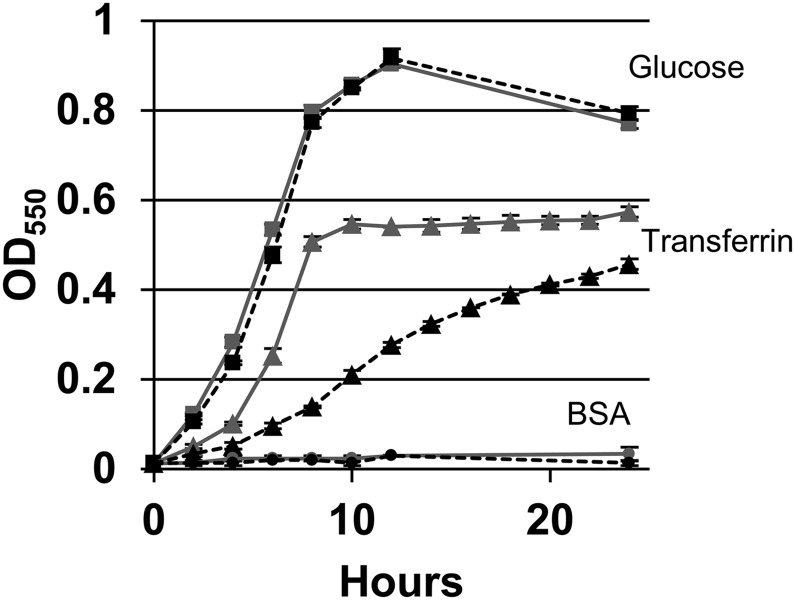
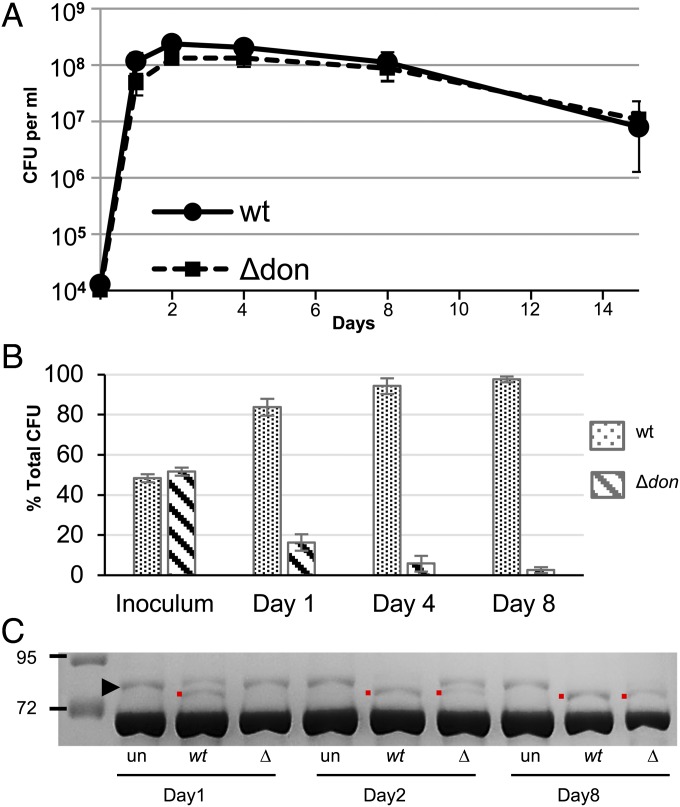
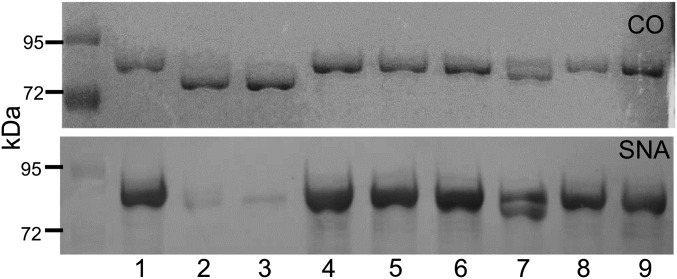
Bacteroides fragilis is the most common anaerobe isolated from clinical infections, and in this report we demonstrate a characteristic of the species that is critical to their success as an opportunistic pathogen. Among the Bacteroides spp. in the gut, B. fragilis has the unique ability of efficiently harvesting complex N-linked glycans from the glycoproteins common to serum and serous fluid. This activity is mediated by an outer membrane protein complex designated as Don. Using the abundant serum glycoprotein transferrin as a model, it has been shown that B. fragilis alone can rapidly and efficiently deglycosylate this protein in vitro and that transferrin glycans can provide the sole source of carbon and energy for growth in defined media. We then showed that transferrin deglycosylation occurs in vivo when B. fragilis is propagated in the rat tissue cage model of extraintestinal growth, and that this ability provides a competitive advantage in vivo over strains lacking the don locus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
