

High-resolution mapping of chromatin packaging in mouse embryonic stem cells and sperm
- PMID: 24998598
- PMCID: PMC4184102
- DOI: 10.1016/j.devcel.2014.05.024
High-resolution mapping of chromatin packaging in mouse embryonic stem cells and sperm
Abstract
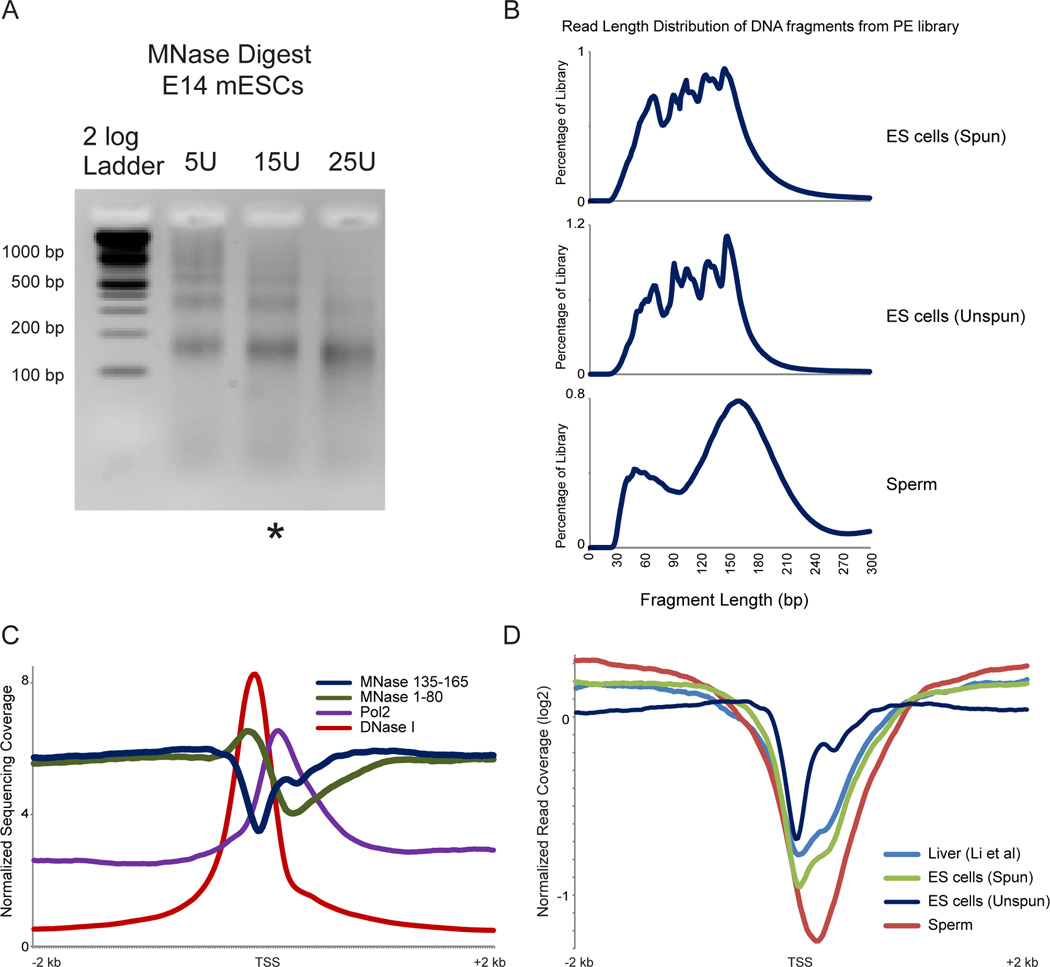
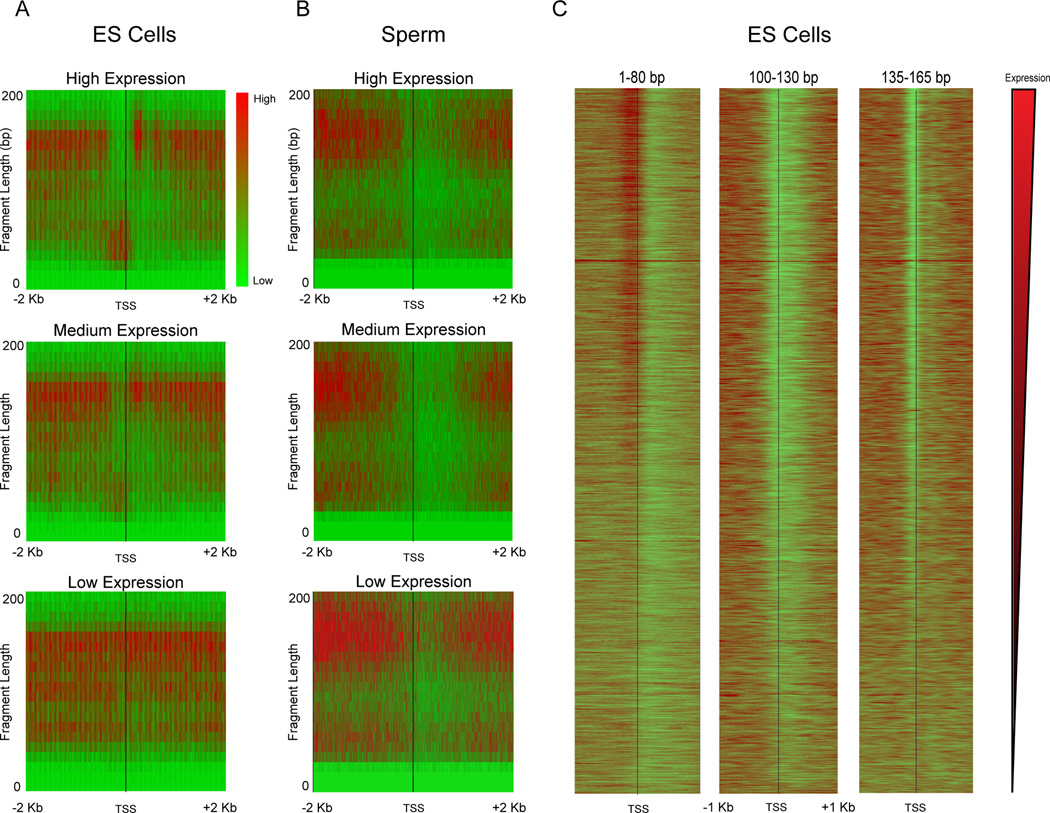
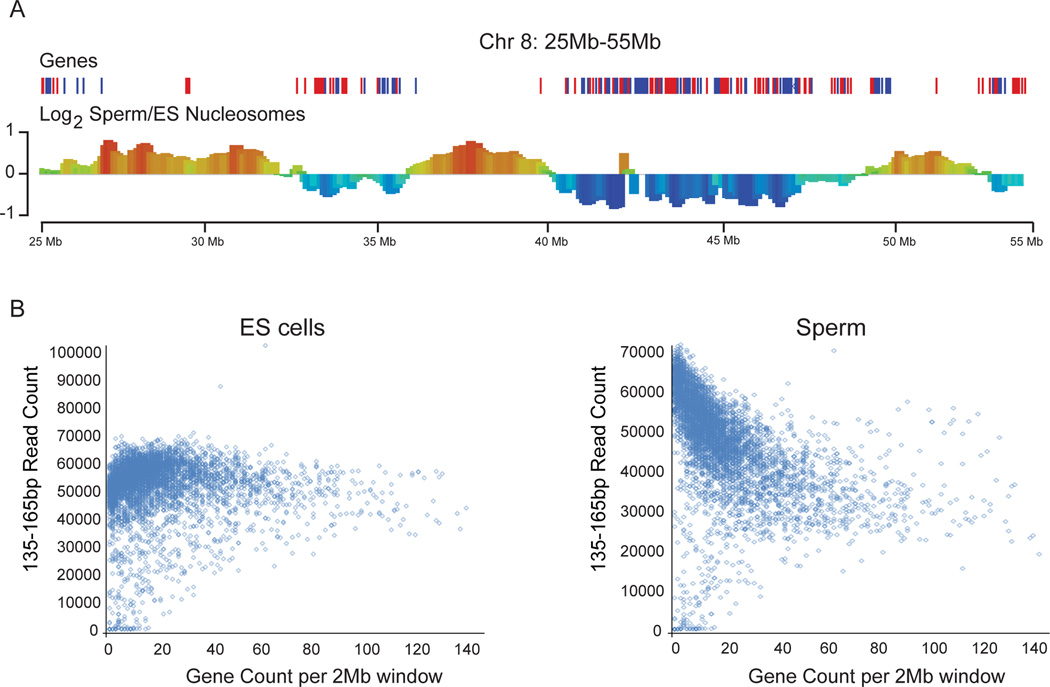
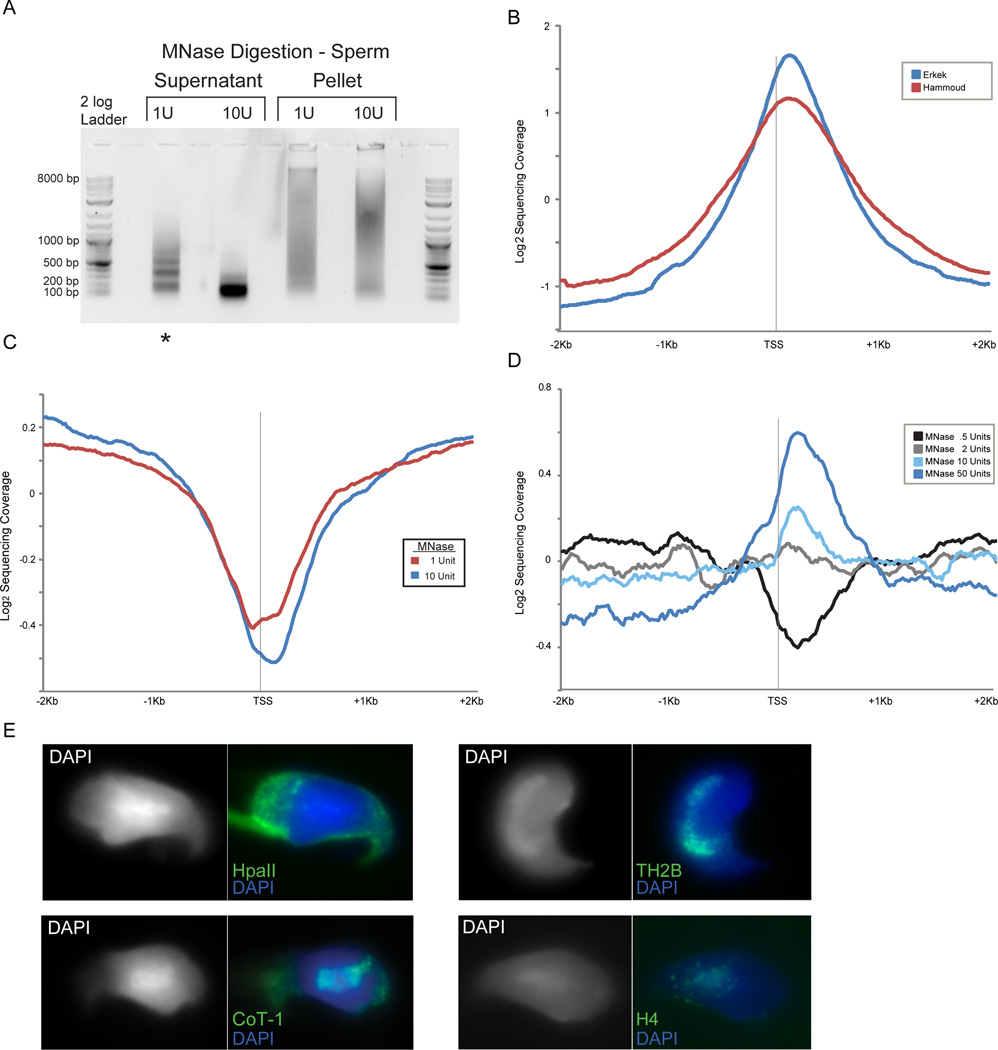
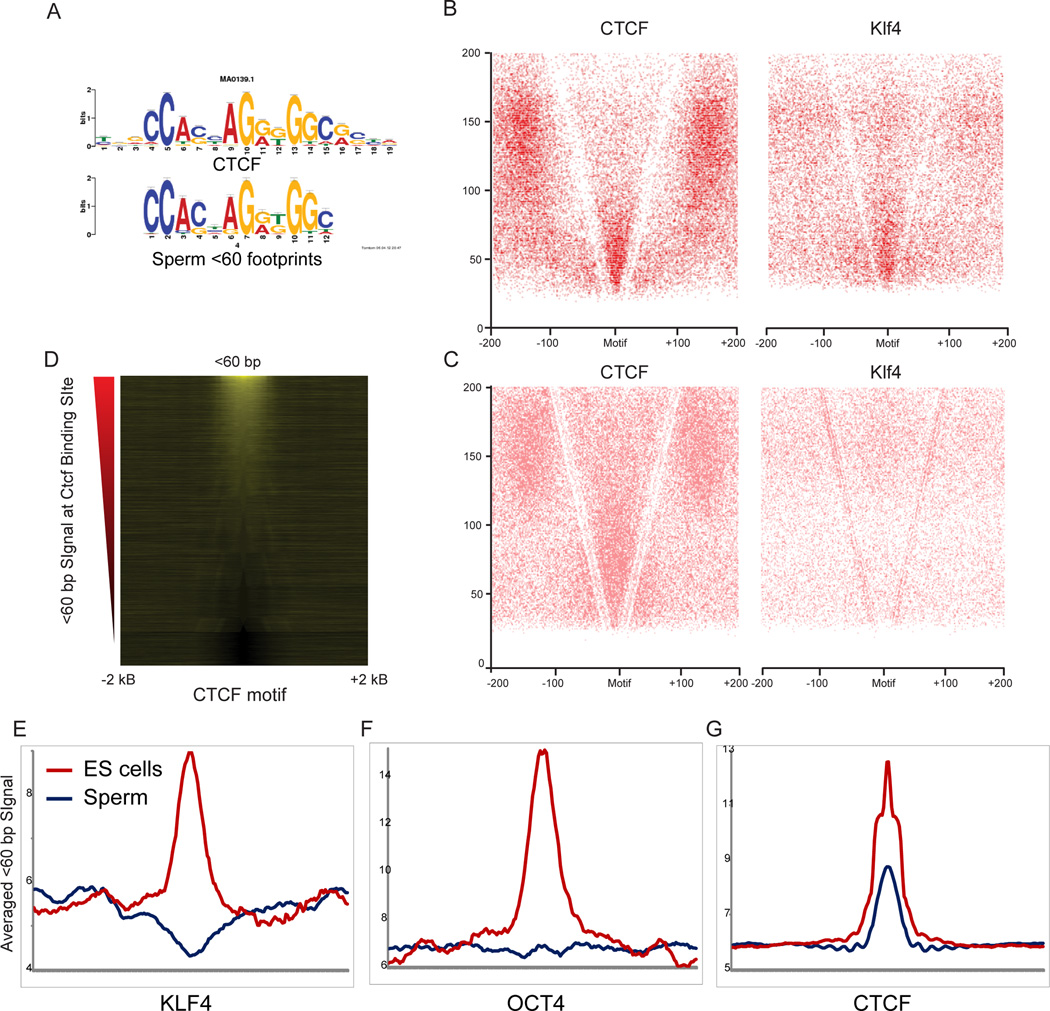
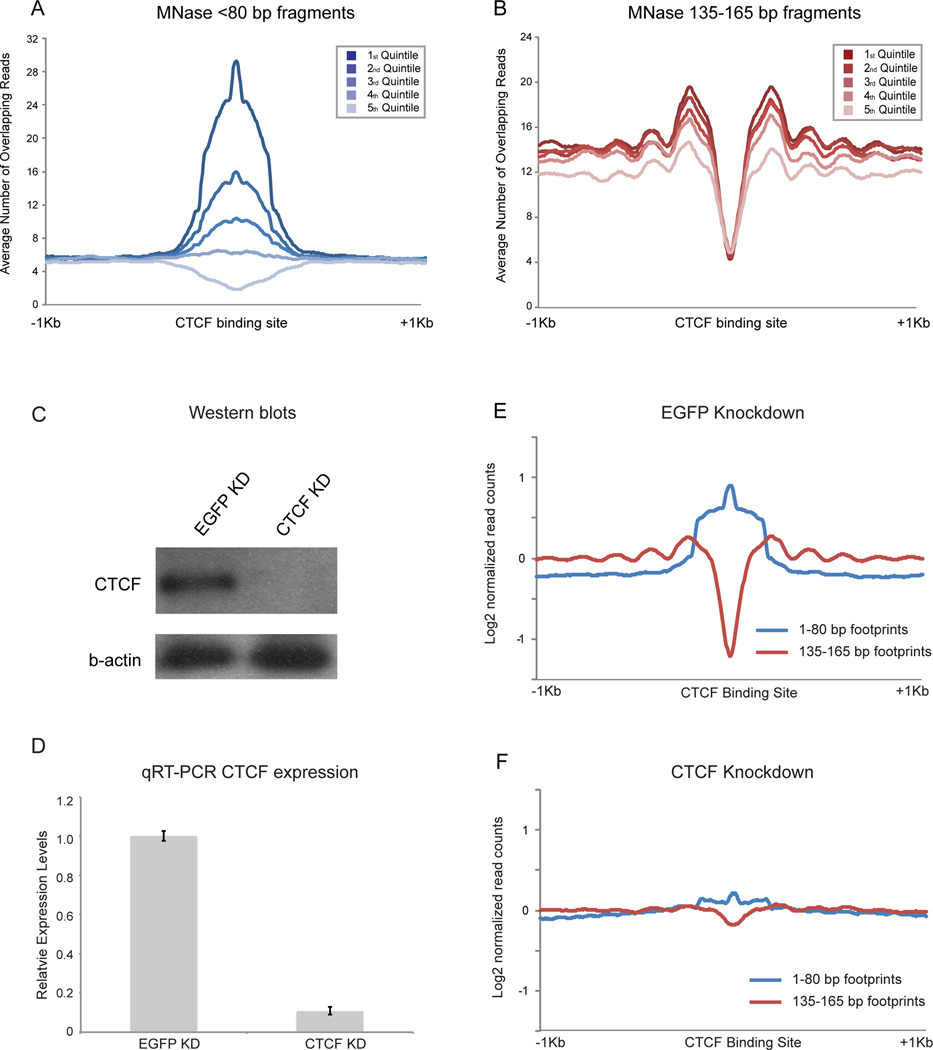
Mammalian embryonic stem cells (ESCs) and sperm exhibit unusual chromatin packaging that plays important roles in cellular function. Here, we extend a recently developed technique, based on deep paired-end sequencing of lightly digested chromatin, to assess footprints of nucleosomes and other DNA-binding proteins genome-wide in murine ESCs and sperm. In ESCs, we recover well-characterized features of chromatin such as promoter nucleosome depletion and further identify widespread footprints of sequence-specific DNA-binding proteins such as CTCF, which we validate in knockdown studies. We document global differences in nuclease accessibility between ESCs and sperm, finding that the majority of histone retention in sperm preferentially occurs in large gene-poor genomic regions, with only a small subset of nucleosomes being retained over promoters of developmental regulators. Finally, we describe evidence that CTCF remains associated with the genome in mature sperm, where it could play a role in organizing the sperm genome.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Paternal nucleosomes: are they retained in developmental promoters or gene deserts?Dev Cell. 2014 Jul 14;30(1):6-8. doi: 10.1016/j.devcel.2014.06.025. Dev Cell. 2014. PMID: 25026032
References
-
- Brykczynska U, Hisano M, Erkek S, Ramos L, Oakeley EJ, Roloff TC, Beisel C, Schubeler D, Stadler MB, Peters AH. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nature structural & molecular biology. 2010;17:679–687. - PubMed
-
- Chen X, Xu H, Yuan P, Fang F, Huss M, Vega VB, Wong E, Orlov YL, Zhang W, Jiang J, et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell. 2008;133:1106–1117. - PubMed
-
- Crick FH, Klug A. Kinky helix. Nature. 1975;255:530–533. - PubMed
-
- Erkek S, Hisano M, Liang CY, Gill M, Murr R, Dieker J, Schubeler D, van der Vlag J, Stadler MB, Peters AH. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nature structural & molecular biology. 2013;20:868–875. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
