

Dependence of brown adipose tissue function on CD36-mediated coenzyme Q uptake
- PMID: 25620701
- PMCID: PMC4318762
- DOI: 10.1016/j.celrep.2014.12.048
Dependence of brown adipose tissue function on CD36-mediated coenzyme Q uptake
Abstract
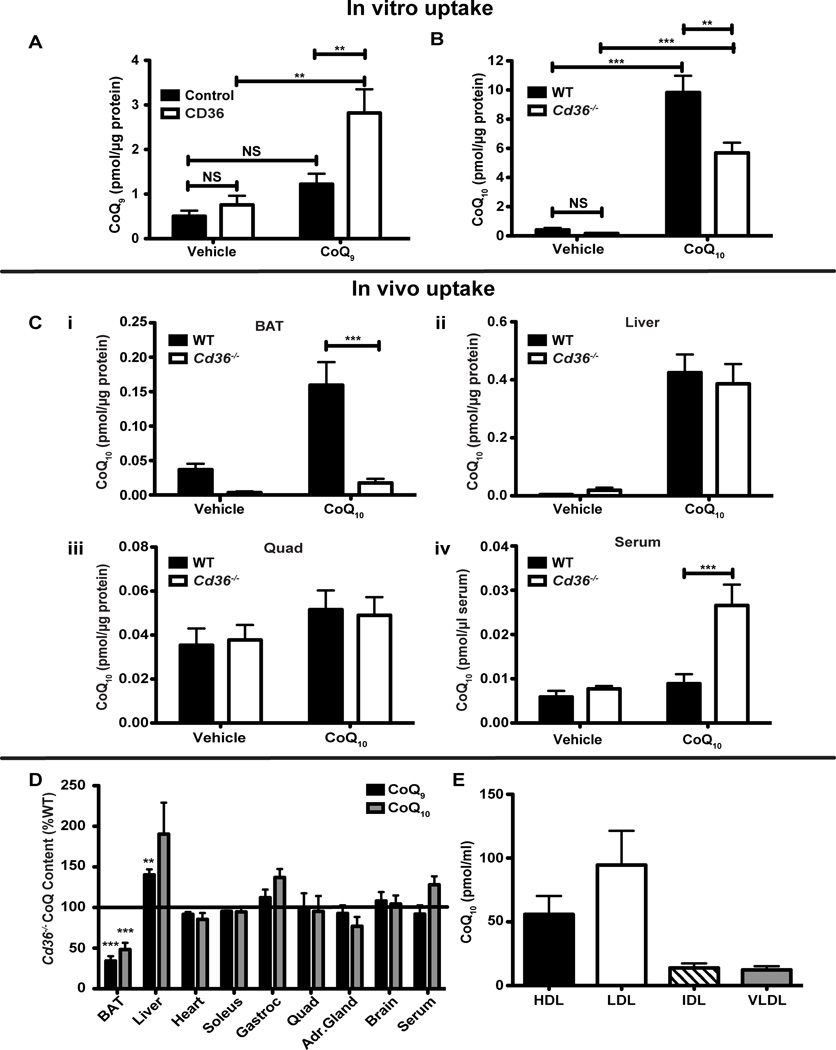
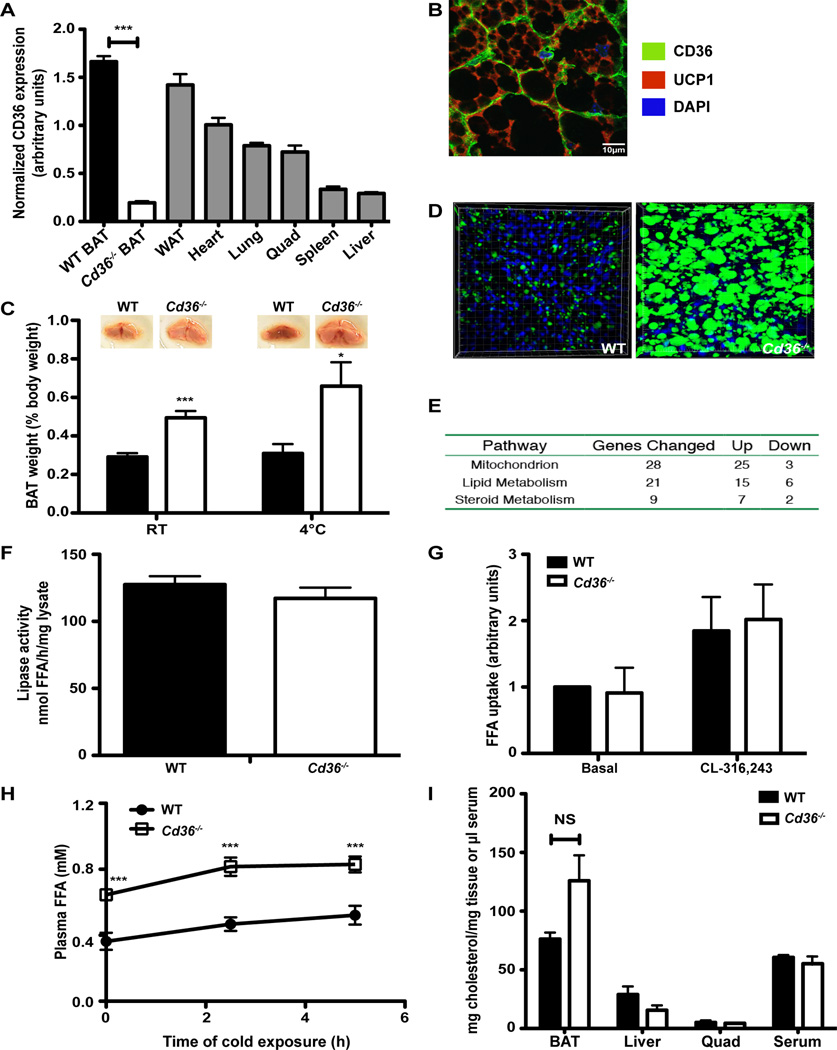
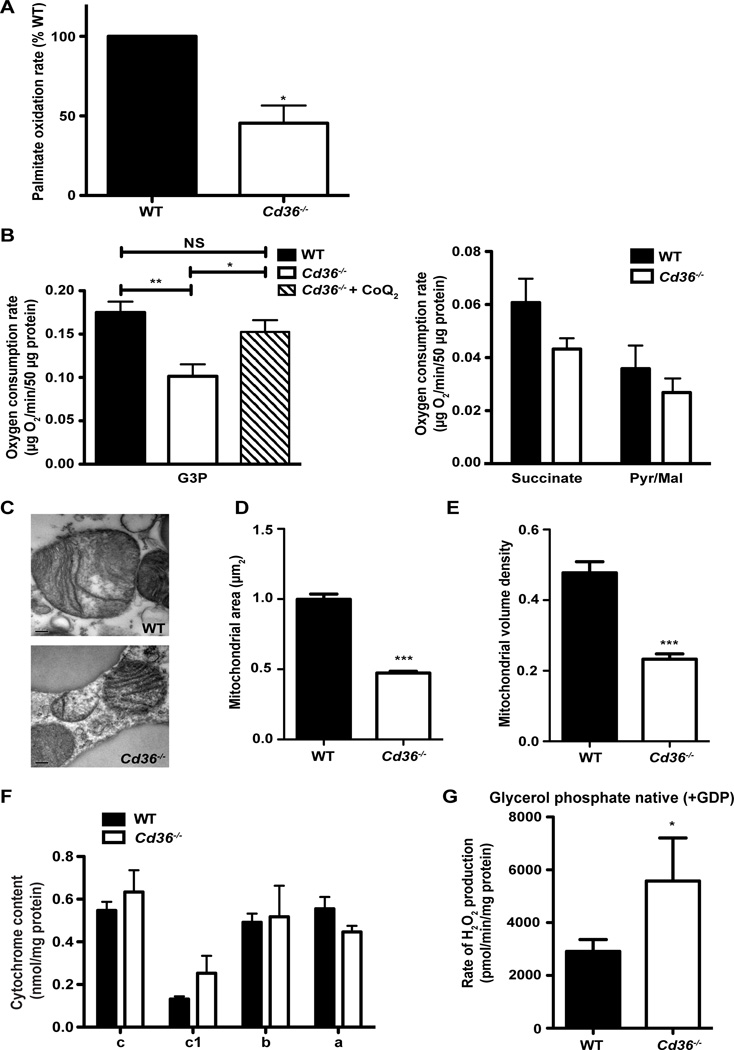
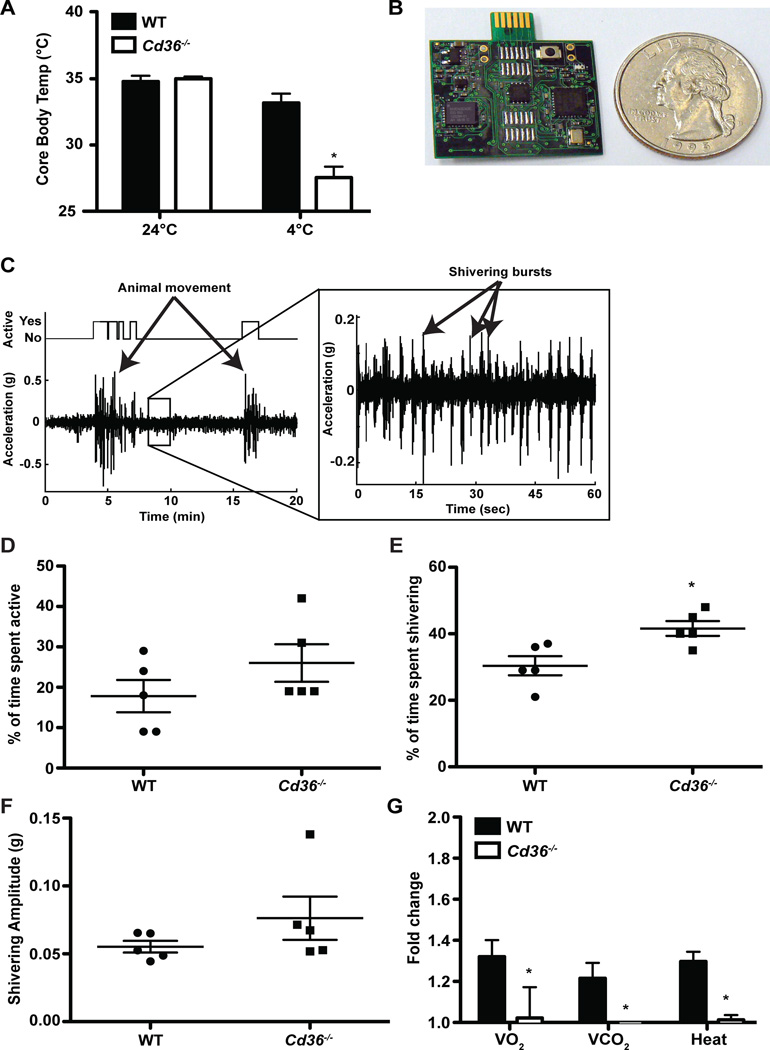
Brown adipose tissue (BAT) possesses the inherent ability to dissipate metabolic energy as heat through uncoupled mitochondrial respiration. An essential component of the mitochondrial electron transport chain is coenzyme Q (CoQ). While cells synthesize CoQ mostly endogenously, exogenous supplementation with CoQ has been successful as a therapy for patients with CoQ deficiency. However, which tissues depend on exogenous CoQ uptake as well as the mechanism by which CoQ is taken up by cells and the role of this process in BAT function are not well understood. Here, we report that the scavenger receptor CD36 drives the uptake of CoQ by BAT and is required for normal BAT function. BAT from mice lacking CD36 displays CoQ deficiency, impaired CoQ uptake, hypertrophy, altered lipid metabolism, mitochondrial dysfunction, and defective nonshivering thermogenesis. Together, these data reveal an important new role for the systemic transport of CoQ to BAT and its function in thermogenesis.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Abumrad NA, el-Maghrabi MR, Amri EZ, Lopez E, Grimaldi PA. Cloning of a rat adipocyte membrane protein implicated in binding or transport of long-chain fatty acids that is induced during preadipocyte differentiation. Homology with human CD36. J Biol Chem. 1993;268:17665–17668. - PubMed
-
- Acton S, Rigotti A, Landschulz KT, Xu S, Hobbs HH, Krieger M. Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science. 1996;271:518–520. - PubMed
-
- Aherne W, Hull D. Brown adipose tissue and heat production in the newborn infant. J Pathol Bacteriol. 1966;91:223–234. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
