

Mincle-mediated translational regulation is required for strong nitric oxide production and inflammation resolution
- PMID: 27089465
- PMCID: PMC4837483
- DOI: 10.1038/ncomms11322
Mincle-mediated translational regulation is required for strong nitric oxide production and inflammation resolution
Abstract
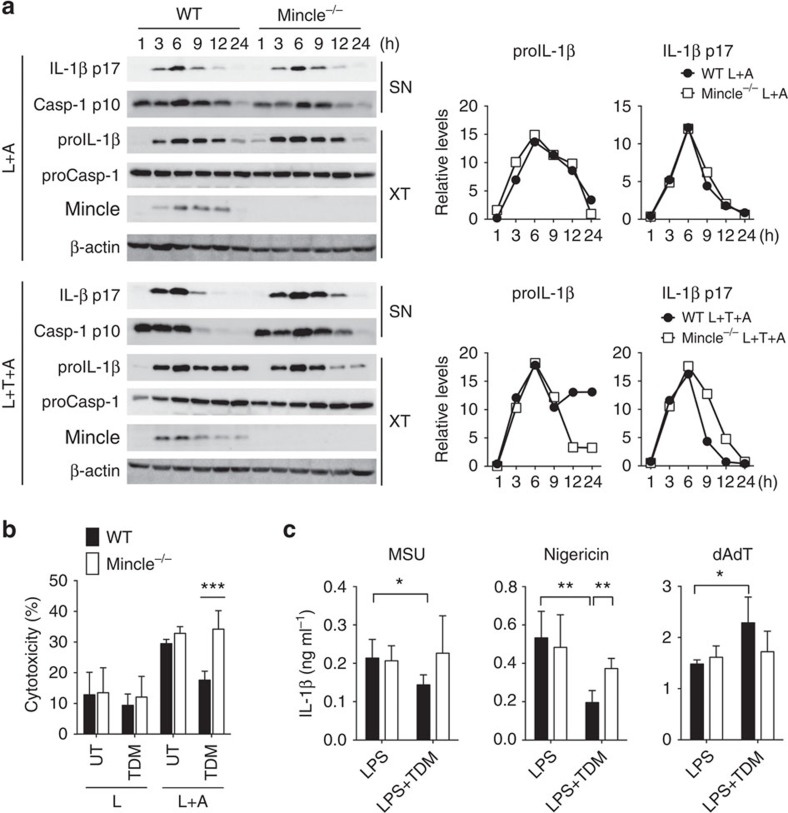
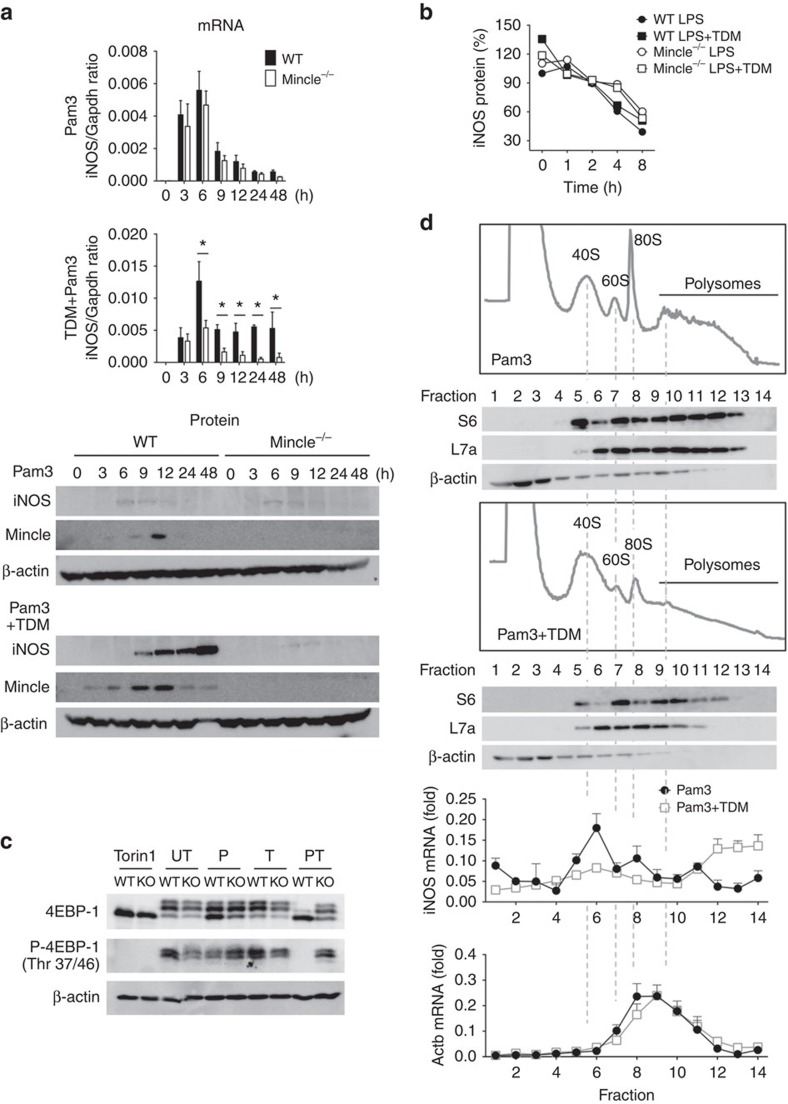
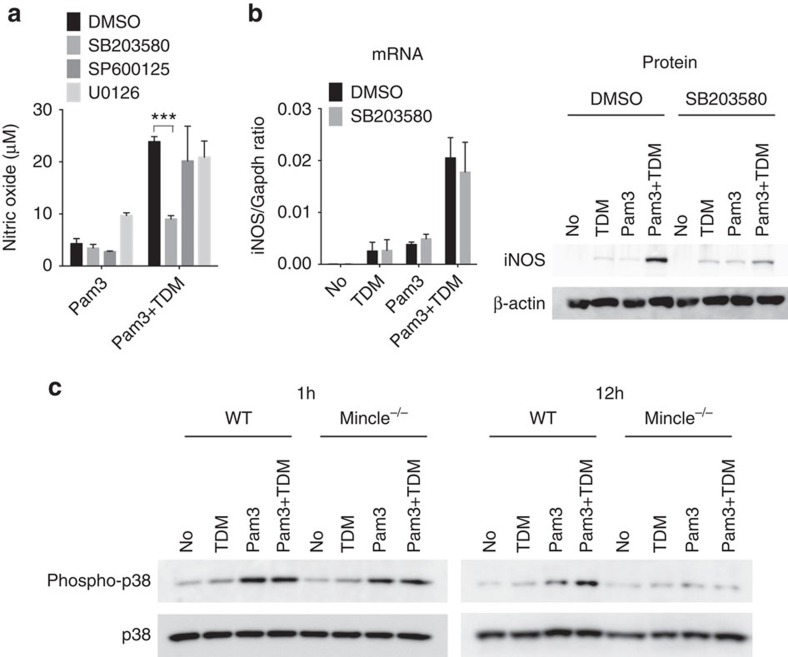
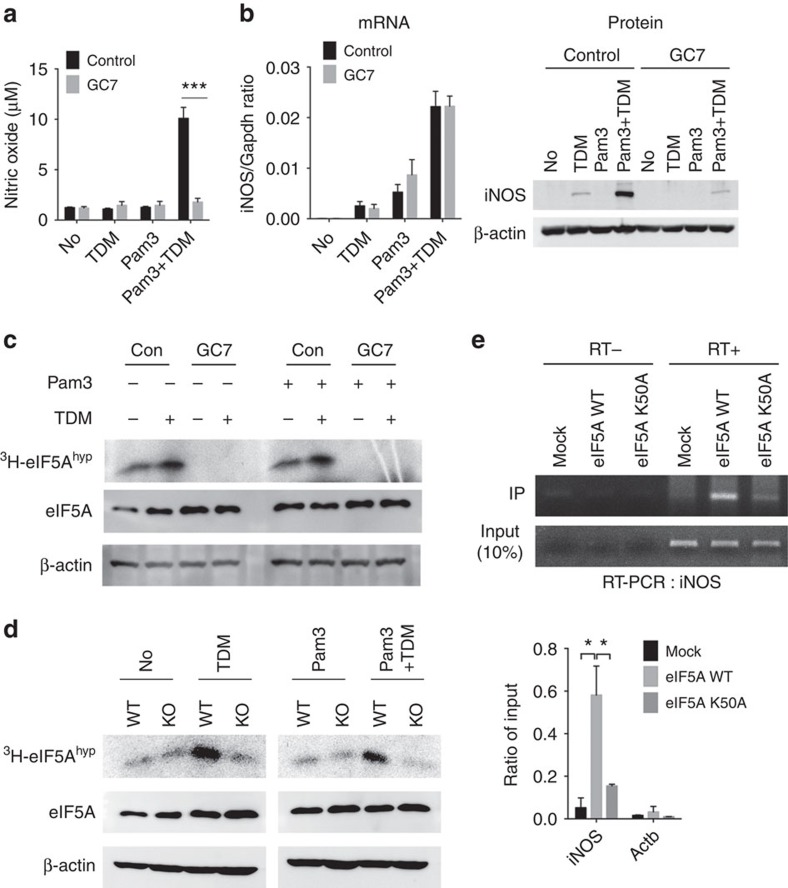
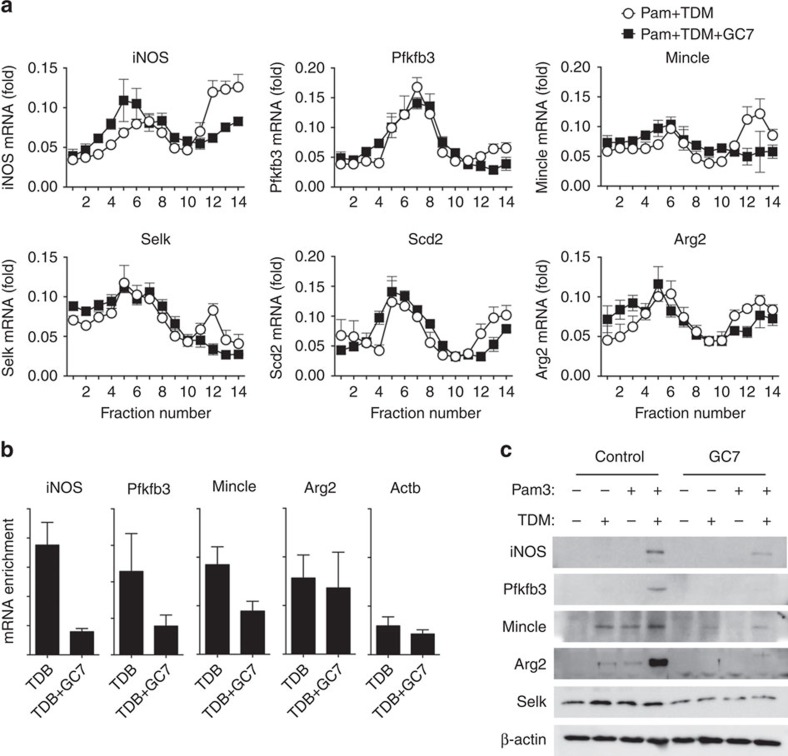
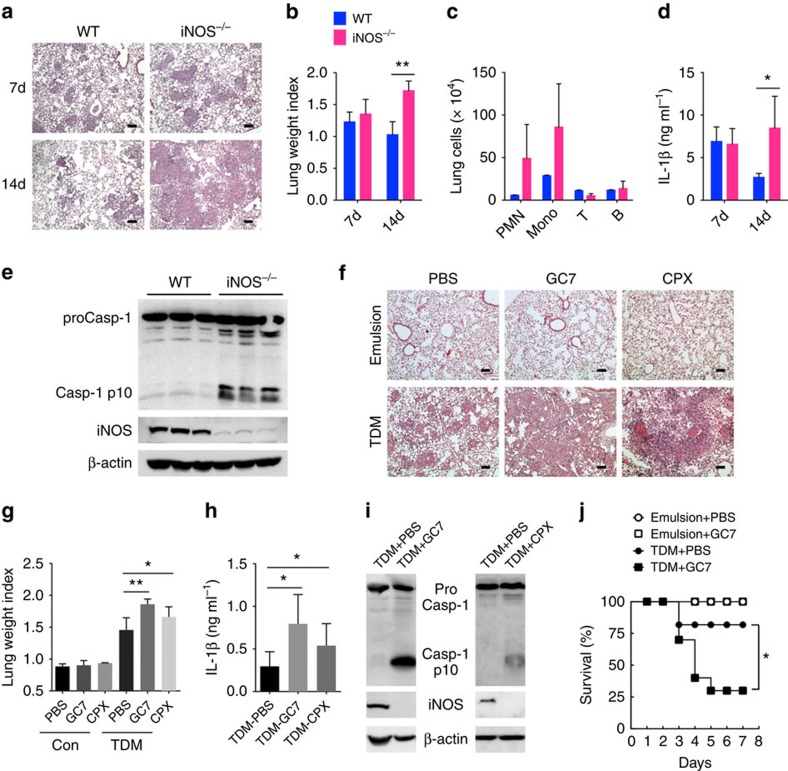
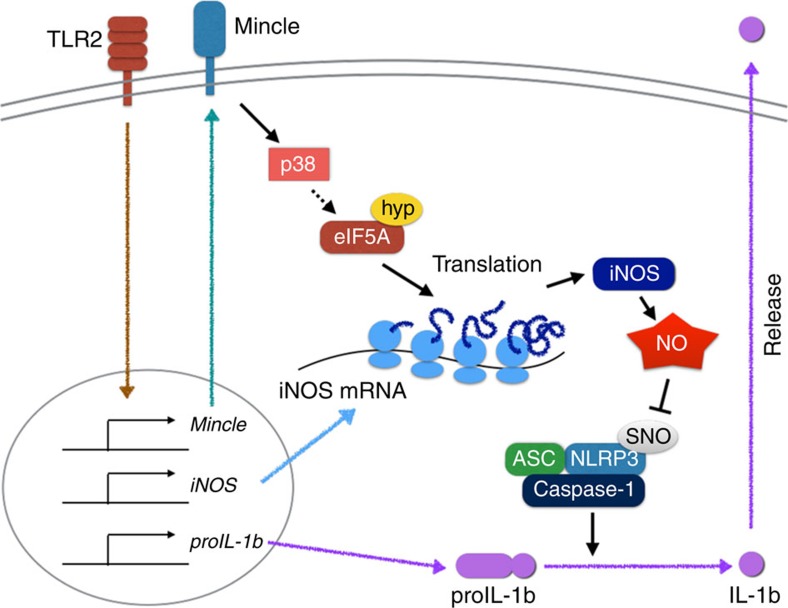
In response to persistent mycobacteria infection, the host induces a granuloma, which often fails to eradicate bacteria and results in tissue damage. Diverse host receptors are required to control the formation and resolution of granuloma, but little is known concerning their regulatory interactions. Here we show that Mincle, the inducible receptor for mycobacterial cord factor, is the key switch for the transition of macrophages from cytokine expression to high nitric oxide production. In addition to its stimulatory role on TLR-mediated transcription, Mincle enhanced the translation of key genes required for nitric oxide synthesis through p38 and eIF5A hypusination, leading to granuloma resolution. Thus, Mincle has dual functions in the promotion and subsequent resolution of inflammation during anti-mycobacterial defence using both transcriptional and translational controls.
Figures

References
-
- Ernst J. D. The immunological life cycle of tuberculosis. Nat. Rev. Immunol. 12, 581–591 (2012). - PubMed
-
- van der Wel N. et al. M. tuberculosis and M. leprae translocate from the phagolysosome to the cytosol in myeloid cells. Cell 129, 1287–1298 (2007). - PubMed
-
- Cambier C. J., Falkow S. & Ramakrishnan L. Host evasion and exploitation schemes of Mycobacterium tuberculosis. Cell 159, 1497–1509 (2014). - PubMed
-
- Ramakrishnan L. Revisiting the role of the granuloma in tuberculosis. Nature 12, 352–366 (2012). - PubMed
-
- Orme I. M., Robinson R. T. & Cooper A. M. The balance between protective and pathogenic immune responses in the TB-infected lung. Nat. Immunol. 16, 57–63 (2015). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
