

Evidence that chytrids dominate fungal communities in high-elevation soils
- PMID: 19826082
- PMCID: PMC2775327
- DOI: 10.1073/pnas.0907303106
Evidence that chytrids dominate fungal communities in high-elevation soils
Abstract
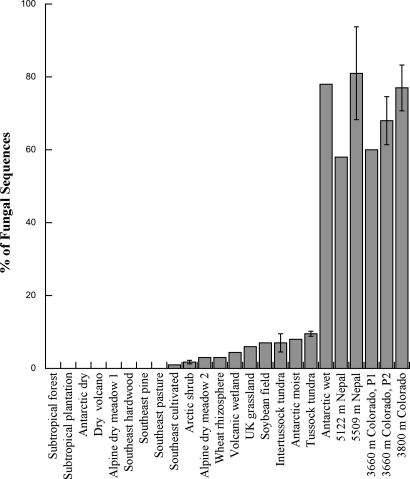
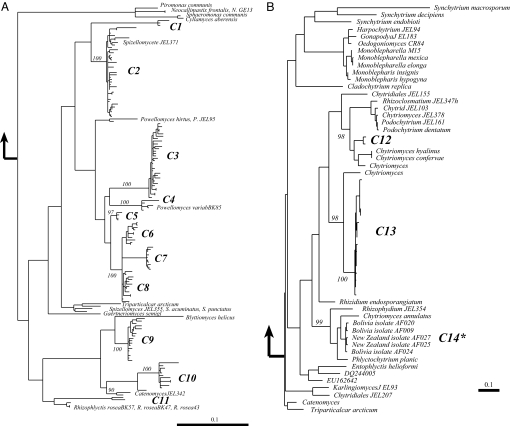
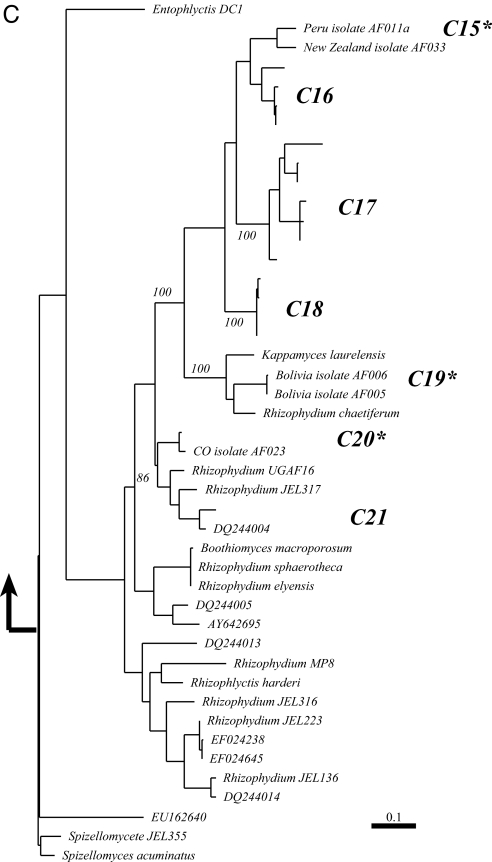
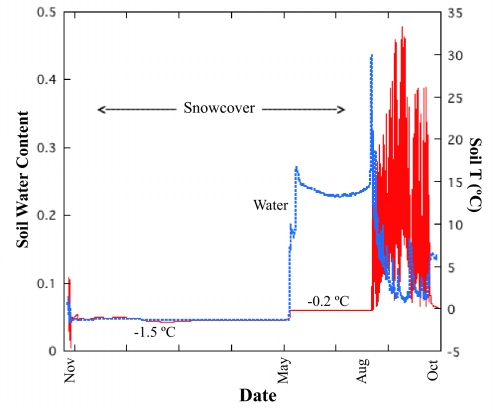
Periglacial soils are one of the least studied ecosystems on Earth, yet they are widespread and are increasing in area due to retreat of glaciers worldwide. Soils in these environments are cold and during the brief summer are exposed to high levels of UV radiation and dramatic fluctuations in moisture and temperature. Recent research suggests that these environments harbor immense microbial diversity. Here we use sequencing of environmental DNA, culturing of isolates, and analysis of environmental variables to show that members of the Chytridiomycota (chytrids) dominate fungal biodiversity and perhaps decomposition processes in plant-free, high-elevation soils from the highest mountain ranges on Earth. The zoosporic reproduction of chytrids requires free water, yet we found that chytrids constituted over 70% of the ribosomal gene sequences of clone libraries from barren soils of the Himalayas and Rockies; by contrast, they are rare in other soil environments. Very few chytrids have been cultured, although we were successful at culturing chytrids from high-elevation sites throughout the world. In a more focused study of our sites in Colorado, we show that carbon sources that support chytrid growth (eolian deposited pollen and microbial phototrophs) are abundant and that soils are saturated with water for several months under the snow, thus creating ideal conditions for the development of a chytrid-dominated ecosystem. Our work broadens the known biodiversity of the Chytridomycota, and describes previously unsuspected links between aquatic and terrestrial ecosystems in alpine regions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Aislabie JM, et al. Dominant bacteria in soils of Marble Point and Wright Valley, Victoria Land, Antarctica. Soil Biol Biochem. 2006;38:3041–3056.
-
- Cowan DA, et al. Antarctic Dry Valley mineral soils contain unexpectedly high levels of microbial biomass. Extremophiles. 2002;6:431–436. - PubMed
-
- Parsons AN, et al. Soil carbon dioxide flux in Antarctic Dry Valley ecosystems. Ecosystems. 2004;7:286–295.
-
- Freeman KR, et al. Soil CO2 flux and photoautotrophic community composition in high-elevation, ‘barren’ soil. Environ Microbiol. 2009;11:674–686. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
