

The Q system: a repressible binary system for transgene expression, lineage tracing, and mosaic analysis
- PMID: 20434990
- PMCID: PMC2883883
- DOI: 10.1016/j.cell.2010.02.025
The Q system: a repressible binary system for transgene expression, lineage tracing, and mosaic analysis
Abstract
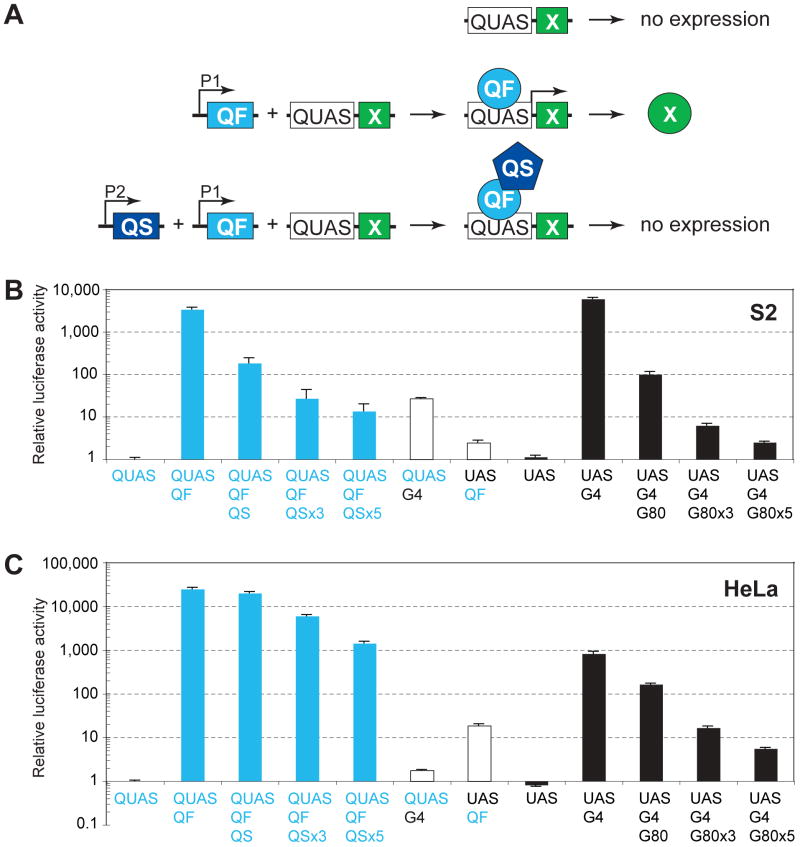
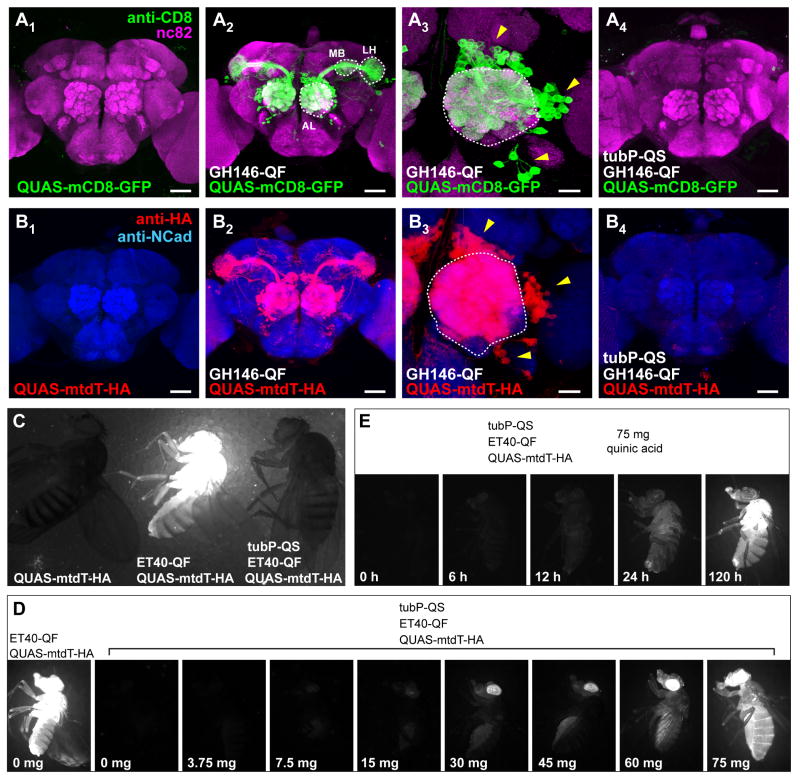
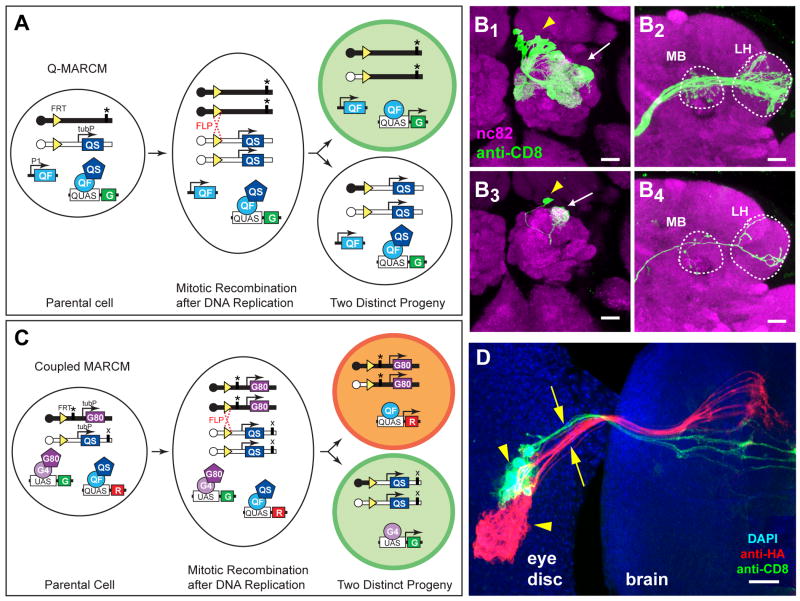
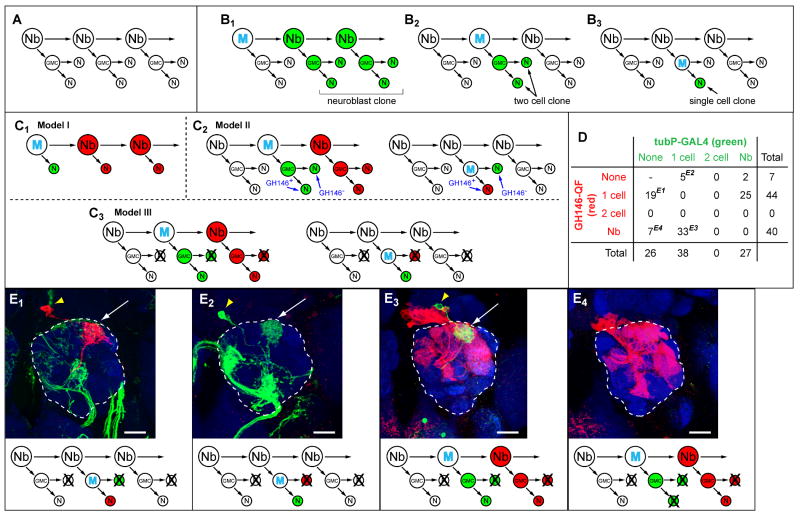
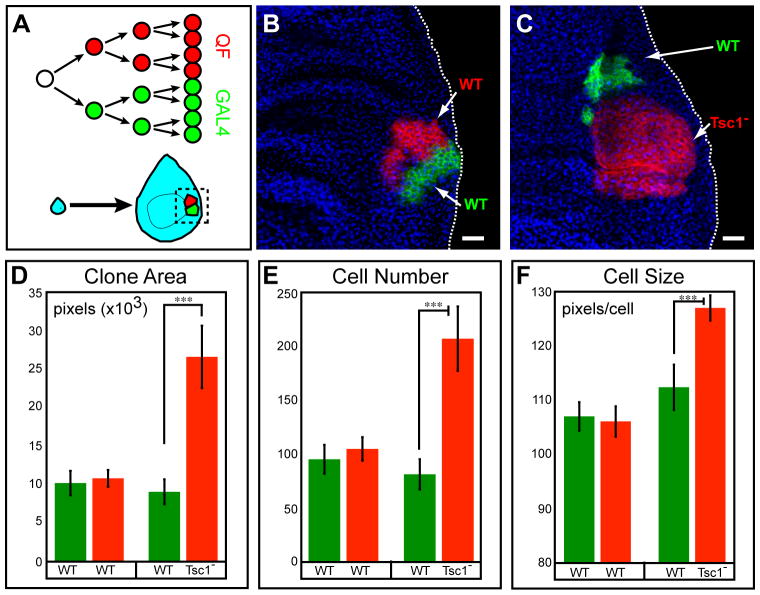
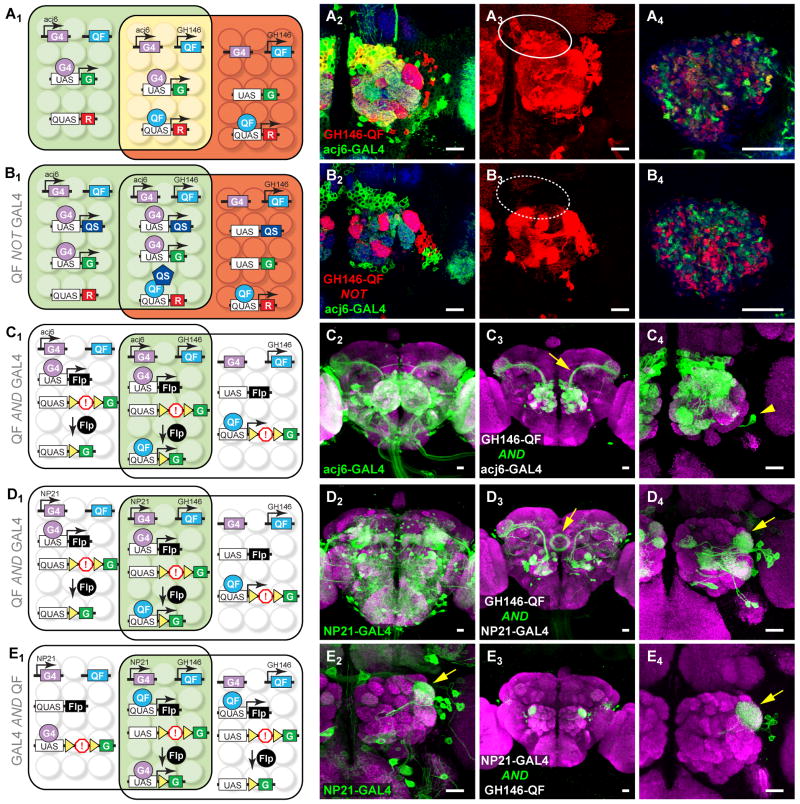
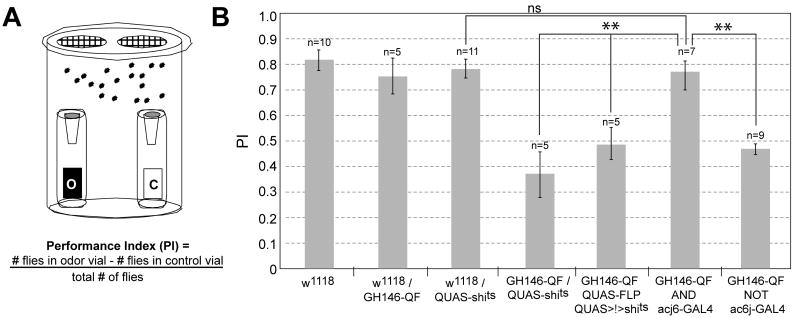
We describe a new repressible binary expression system based on the regulatory genes from the Neurospora qa gene cluster. This "Q system" offers attractive features for transgene expression in Drosophila and mammalian cells: low basal expression in the absence of the transcriptional activator QF, high QF-induced expression, and QF repression by its repressor QS. Additionally, feeding flies quinic acid can relieve QS repression. The Q system offers many applications, including (1) intersectional "logic gates" with the GAL4 system for manipulating transgene expression patterns, (2) GAL4-independent MARCM analysis, and (3) coupled MARCM analysis to independently visualize and genetically manipulate siblings from any cell division. We demonstrate the utility of the Q system in determining cell division patterns of a neuronal lineage and gene function in cell growth and proliferation, and in dissecting neurons responsible for olfactory attraction. The Q system can be expanded to other uses in Drosophila and to any organism conducive to transgenesis.
2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Bryant PJ, Simpson P. Intrinsic and extrinsic control of growth in developing organs. Q Rev Biol. 1984;59:387–415. - PubMed
-
- Clyne PJ, Certel SJ, de Bruyne M, Zaslavsky L, Johnson WA, Carlson JR. The odor specificities of a subset of olfactory receptor neurons are governed by Acj6, a POU-domain transcription factor. Neuron. 1999;22:339–347. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
