

PRDM9 variation strongly influences recombination hot-spot activity and meiotic instability in humans
- PMID: 20818382
- PMCID: PMC3092422
- DOI: 10.1038/ng.658
PRDM9 variation strongly influences recombination hot-spot activity and meiotic instability in humans
Abstract
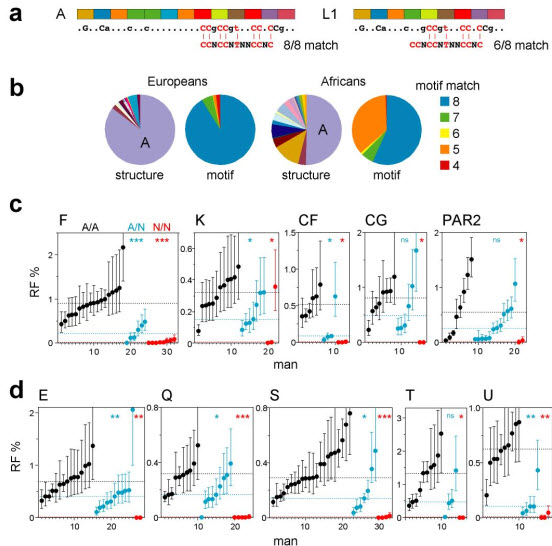
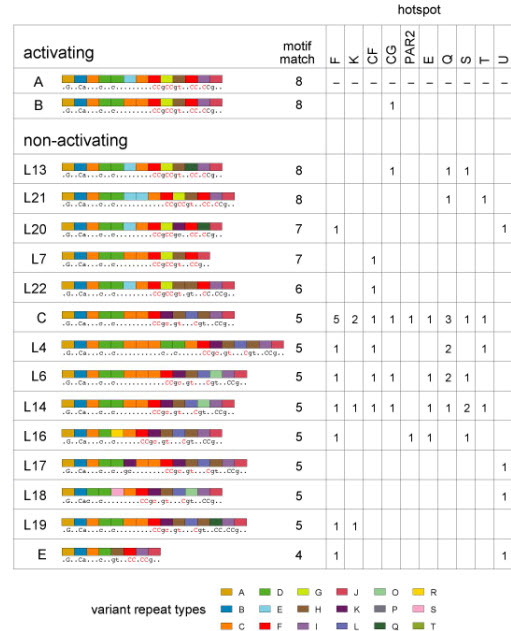
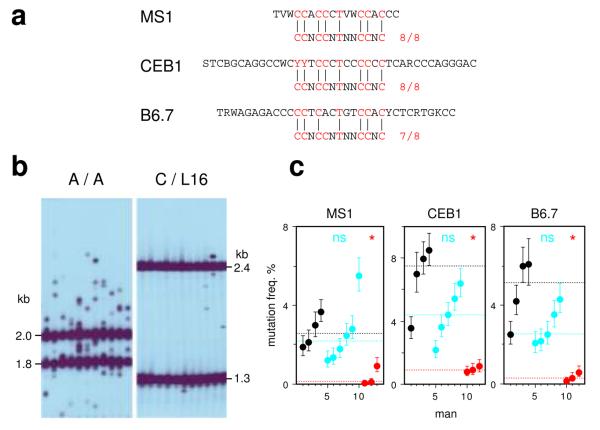
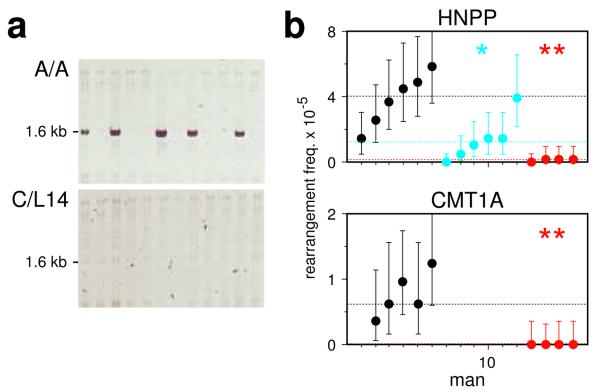
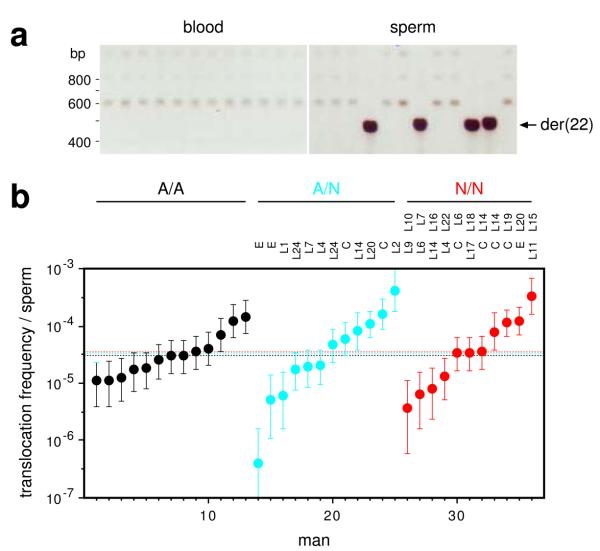
PRDM9 has recently been identified as a likely trans regulator of meiotic recombination hot spots in humans and mice. PRDM9 contains a zinc finger array that, in humans, can recognize a short sequence motif associated with hot spots, with binding to this motif possibly triggering hot-spot activity via chromatin remodeling. We now report that human genetic variation at the PRDM9 locus has a strong effect on sperm hot-spot activity, even at hot spots lacking the sequence motif. Subtle changes within the zinc finger array can create hot-spot nonactivating or enhancing variants and can even trigger the appearance of a new hot spot, suggesting that PRDM9 is a major global regulator of hot spots in humans. Variation at the PRDM9 locus also influences aspects of genome instability-specifically, a megabase-scale rearrangement underlying two genomic disorders as well as minisatellite instability-implicating PRDM9 as a risk factor for some pathological genome rearrangements.
Figures

Comment in
-
PRDM9 marks the spot.Nat Genet. 2010 Oct;42(10):821-2. doi: 10.1038/ng1010-821. Nat Genet. 2010. PMID: 20877321 No abstract available.
References
-
- Myers S, Freeman C, Auton A, Donnelly P, McVean G. A common sequence motif associated with recombination hot spots and genome instability in humans. Nat. Genet. 2008;40:1124–1129. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
