

Repeatless and repeat-based centromeres in potato: implications for centromere evolution
- PMID: 22968715
- PMCID: PMC3480287
- DOI: 10.1105/tpc.112.100511
Repeatless and repeat-based centromeres in potato: implications for centromere evolution
Abstract
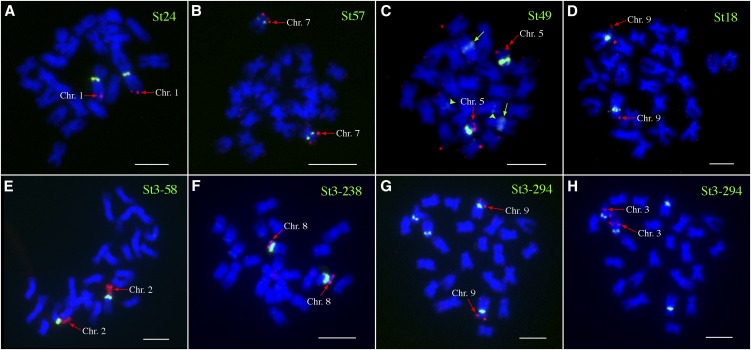
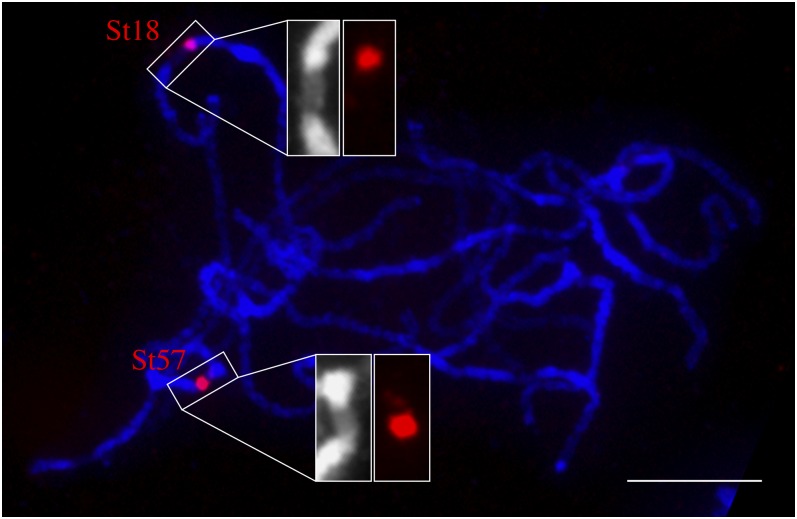
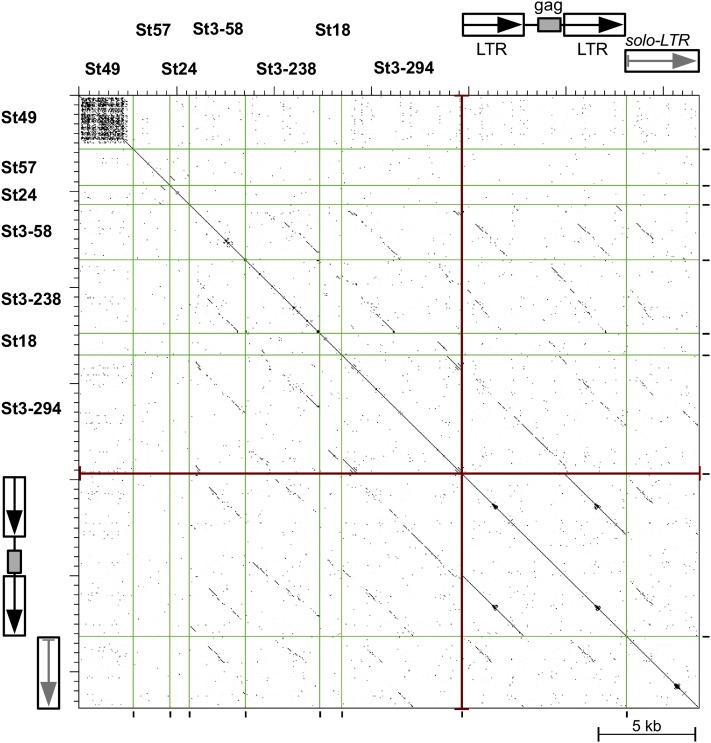
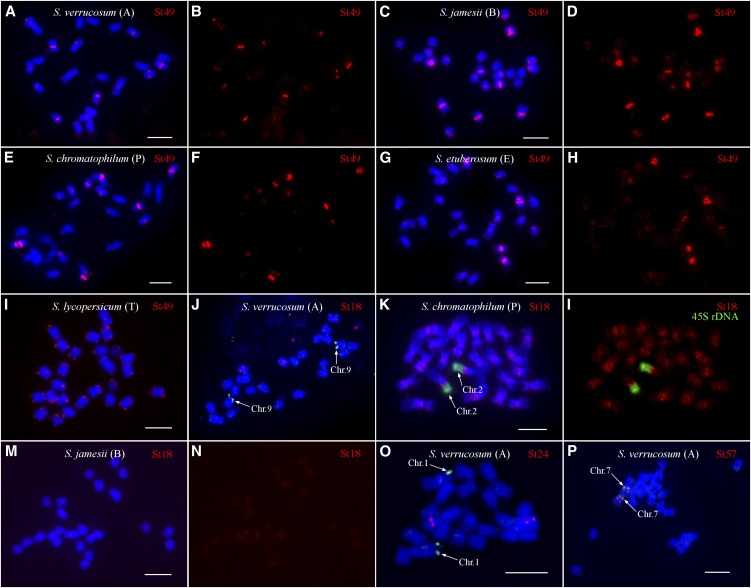
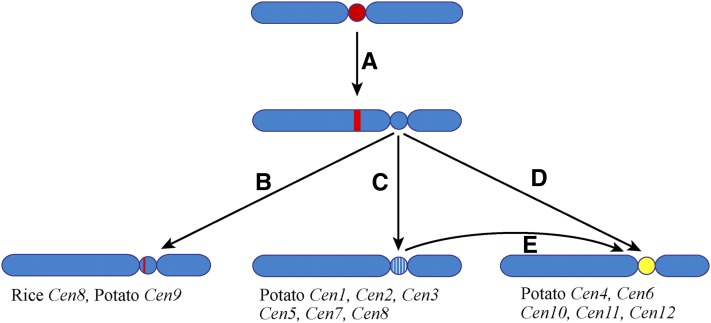
Centromeres in most higher eukaryotes are composed of long arrays of satellite repeats. By contrast, most newly formed centromeres (neocentromeres) do not contain satellite repeats and instead include DNA sequences representative of the genome. An unknown question in centromere evolution is how satellite repeat-based centromeres evolve from neocentromeres. We conducted a genome-wide characterization of sequences associated with CENH3 nucleosomes in potato (Solanum tuberosum). Five potato centromeres (Cen4, Cen6, Cen10, Cen11, and Cen12) consisted primarily of single- or low-copy DNA sequences. No satellite repeats were identified in these five centromeres. At least one transcribed gene was associated with CENH3 nucleosomes. Thus, these five centromeres structurally resemble neocentromeres. By contrast, six potato centromeres (Cen1, Cen2, Cen3, Cen5, Cen7, and Cen8) contained megabase-sized satellite repeat arrays that are unique to individual centromeres. The satellite repeat arrays likely span the entire functional cores of these six centromeres. At least four of the centromeric repeats were amplified from retrotransposon-related sequences and were not detected in Solanum species closely related to potato. The presence of two distinct types of centromeres, coupled with the boom-and-bust cycles of centromeric satellite repeats in Solanum species, suggests that repeat-based centromeres can rapidly evolve from neocentromeres by de novo amplification and insertion of satellite repeats in the CENH3 domains.
Figures
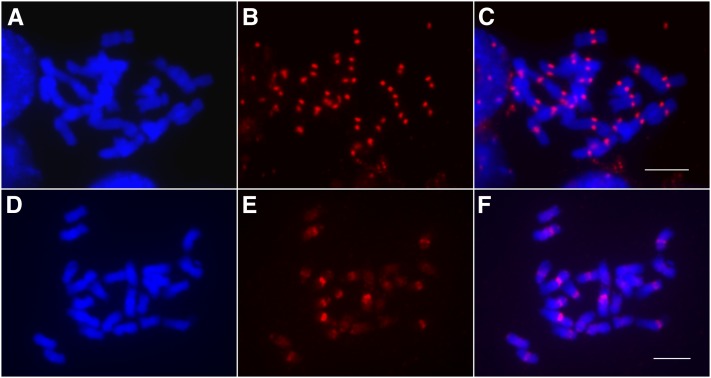
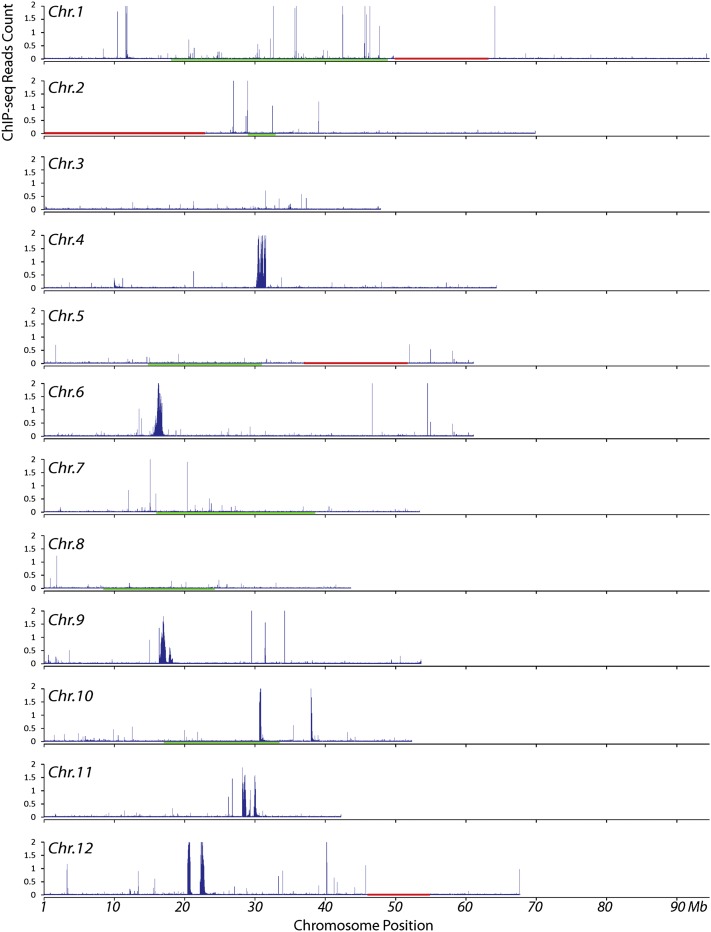
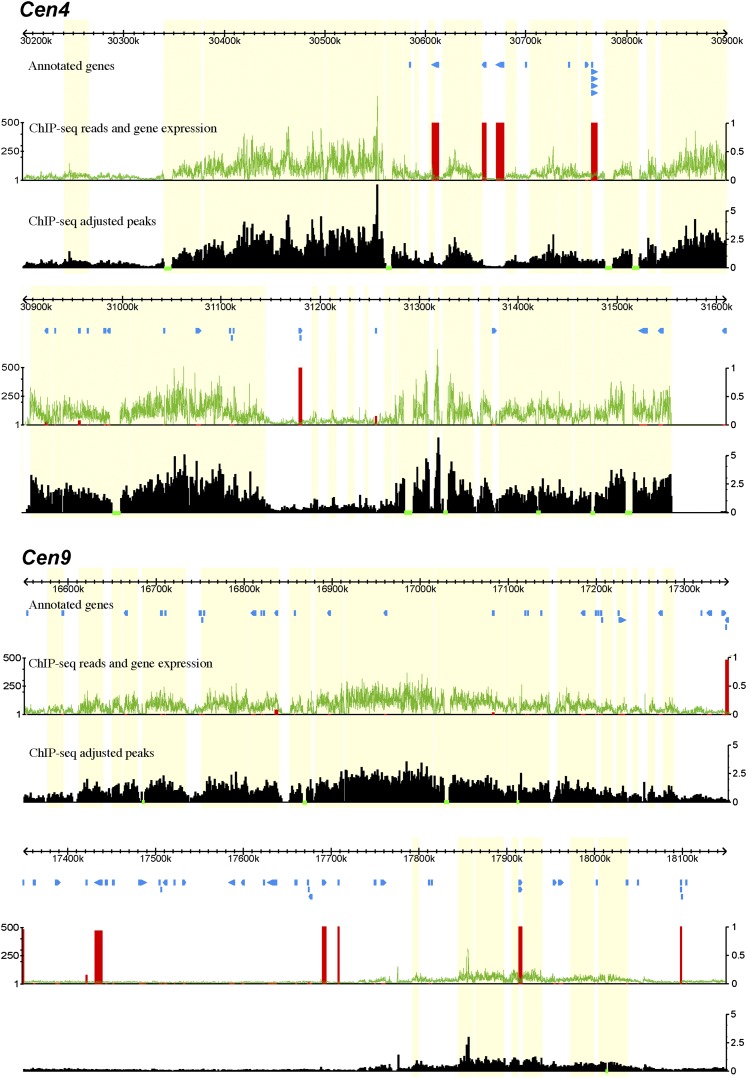
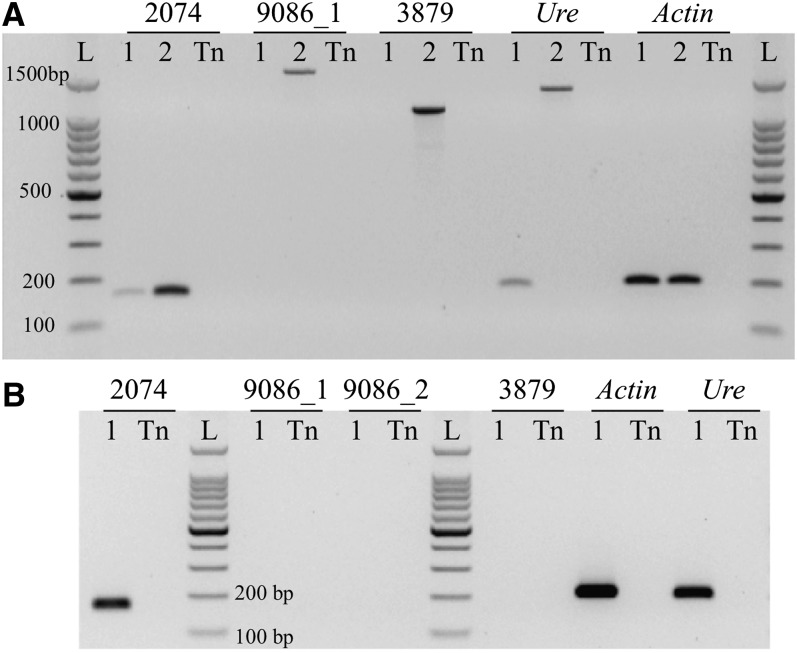

Comment in
-
Rapid centromere evolution in potato: invasion of the satellite repeats.Plant Cell. 2012 Sep;24(9):3487. doi: 10.1105/tpc.112.240912. Epub 2012 Sep 25. Plant Cell. 2012. PMID: 23012435 Free PMC article. No abstract available.
References
-
- Allshire R.C., Nimmo E.R., Ekwall K., Javerzat J.P., Cranston G. (1995). Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev. 9: 218–233 - PubMed
-
- Castillo R.O., Spooner D.M. (1997). Phylogenetic relationships of wild potatoes, Solanum series Conicibaccata (sect. Petota). Syst. Bot. 22: 45–83
-
- Cheng Z.K., Buell C.R., Wing R.A., Jiang J. (2002). Resolution of fluorescence in-situ hybridization mapping on rice mitotic prometaphase chromosomes, meiotic pachytene chromosomes and extended DNA fibers. Chromosome Res. 10: 379–387 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
