

Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed
- PMID: 26324896
- PMCID: PMC4577148
- DOI: 10.1073/pnas.1502160112
Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed
Abstract
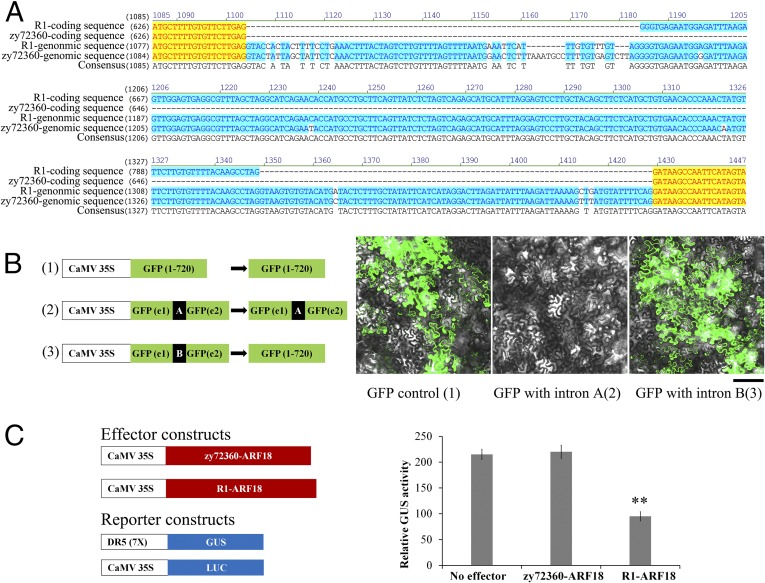
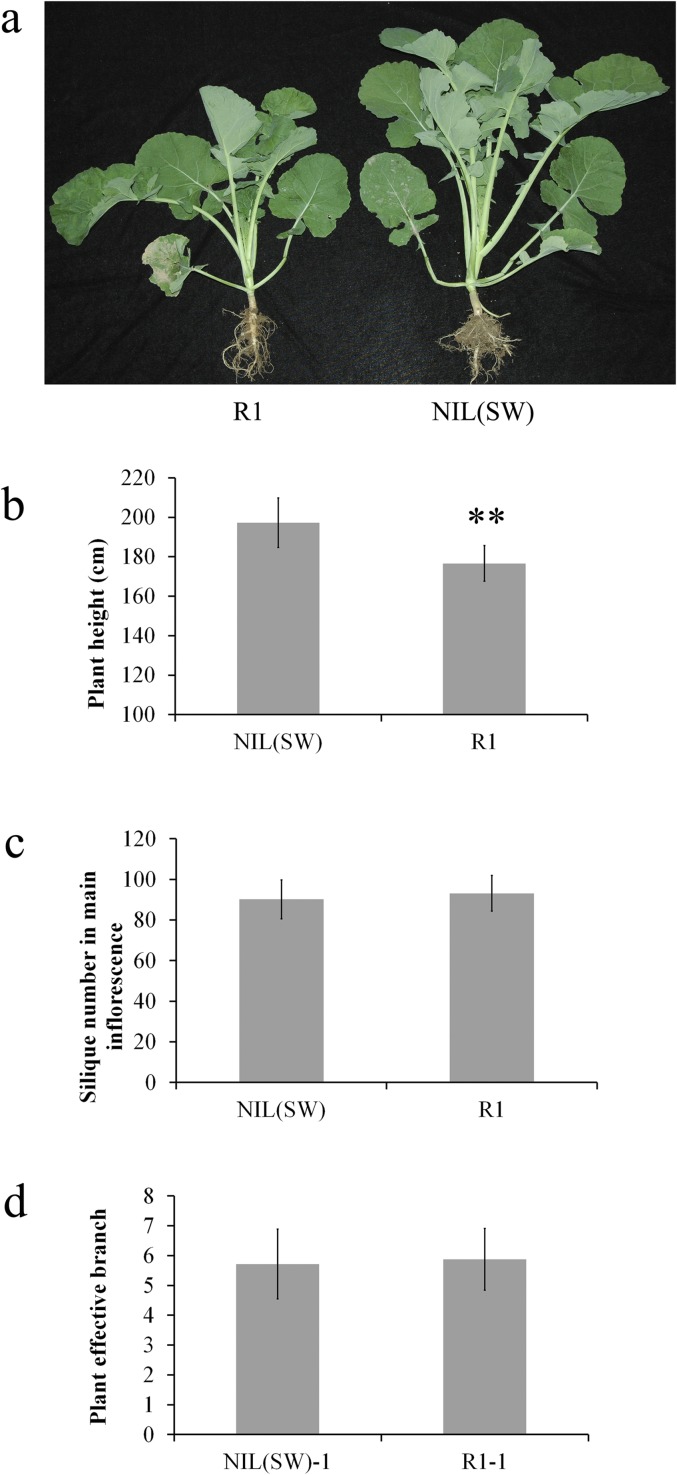
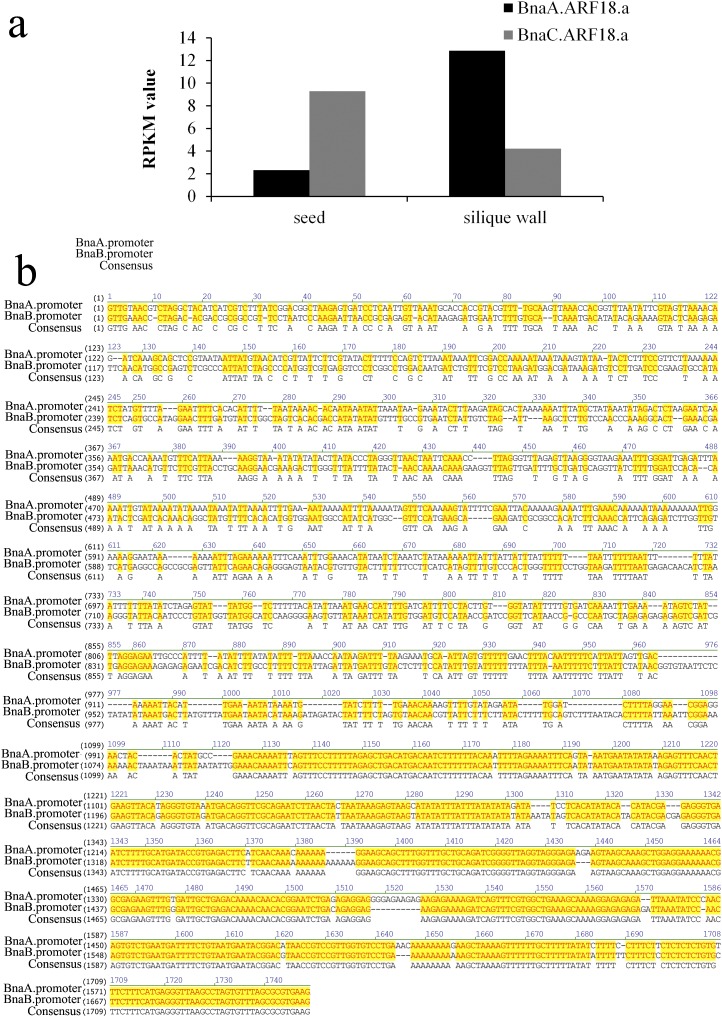
Seed weight (SW), which is one of the three major factors influencing grain yield, has been widely accepted as a complex trait that is controlled by polygenes, particularly in polyploid crops. Brassica napus L., which is the second leading crop source for vegetable oil around the world, is a tetraploid (4×) species. In the present study, we identified a major quantitative trait locus (QTL) on chromosome A9 of rapeseed in which the genes for SW and silique length (SL) were colocated. By fine mapping and association analysis, we uncovered a 165-bp deletion in the auxin-response factor 18 (ARF18) gene associated with increased SW and SL. ARF18 encodes an auxin-response factor and shows inhibitory activity on downstream auxin genes. This 55-aa deletion prevents ARF18 from forming homodimers, in turn resulting in the loss of binding activity. Furthermore, reciprocal crossing has shown that this QTL affects SW by maternal effects. Transcription analysis has shown that ARF18 regulates cell growth in the silique wall by acting via an auxin-response pathway. Together, our results suggest that ARF18 regulates silique wall development and determines SW via maternal regulation. In addition, our study reveals the first (to our knowledge) QTL in rapeseed and may provide insights into gene cloning involving polyploid crops.
Keywords: ARF18; cell growth; maternal effect; seed weight; silique length.
Conflict of interest statement
The authors declare no conflict of interest.
Figures










References
-
- Fang W, Wang Z, Cui R, Li J, Li Y. Maternal control of seed size by EOD3/CYP78A6 in Arabidopsis thaliana. Plant J. 2012;70(6):929–939. - PubMed
-
- Schruff MC, et al. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development. 2006;133(2):251–261. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
