

Recurring exon deletions in the HP (haptoglobin) gene contribute to lower blood cholesterol levels
- PMID: 26901066
- PMCID: PMC4811681
- DOI: 10.1038/ng.3510
Recurring exon deletions in the HP (haptoglobin) gene contribute to lower blood cholesterol levels
Abstract
One of the first protein polymorphisms identified in humans involves the abundant blood protein haptoglobin. Two exons of the HP gene (encoding haptoglobin) exhibit copy number variation that affects HP protein structure and multimerization. The evolutionary origins and medical relevance of this polymorphism have been uncertain. Here we show that this variation has likely arisen from many recurring deletions, more specifically, reversions of an ancient hominin-specific duplication of these exons. Although this polymorphism has been largely invisible to genome-wide genetic studies thus far, we describe a way to analyze it by imputation from SNP haplotypes and find among 22,288 individuals that these HP exonic deletions associate with reduced LDL and total cholesterol levels. We further show that these deletions, and a SNP that affects HP expression, appear to drive the strong association of cholesterol levels with SNPs near HP. Recurring exonic deletions in HP likely enhance human health by lowering cholesterol levels in the blood.
Figures
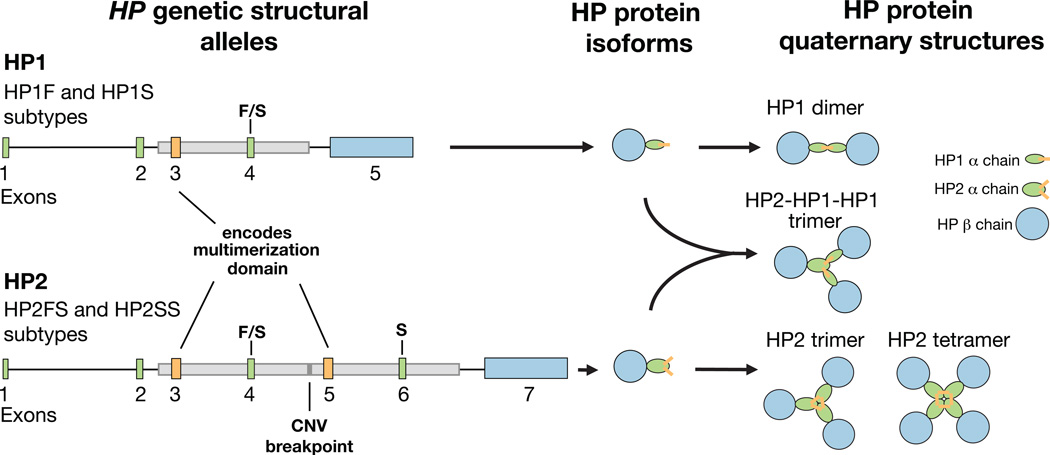
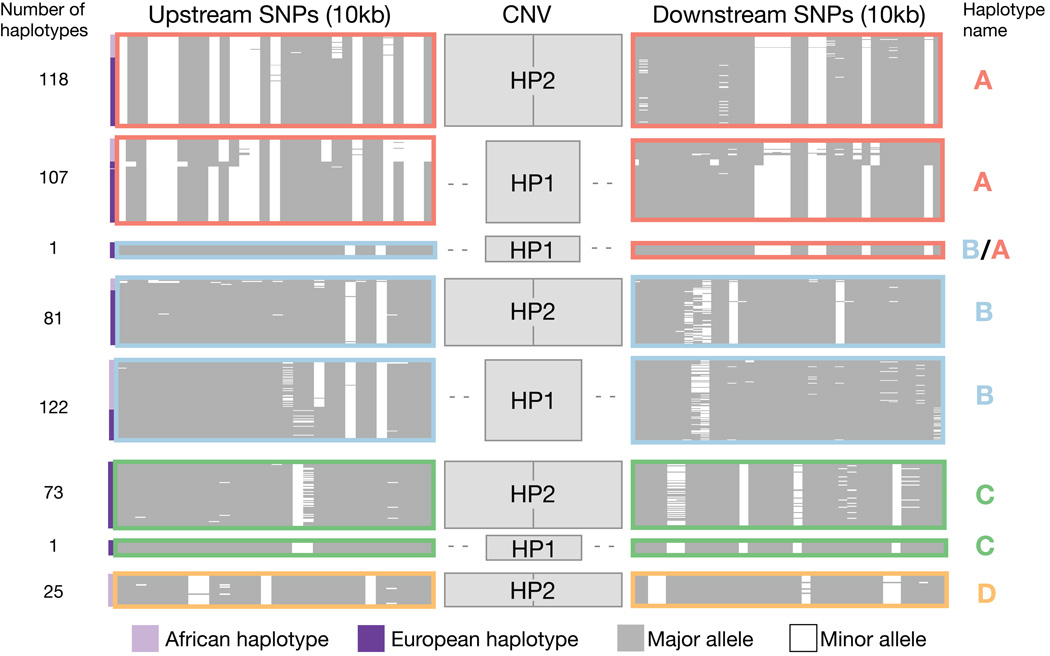
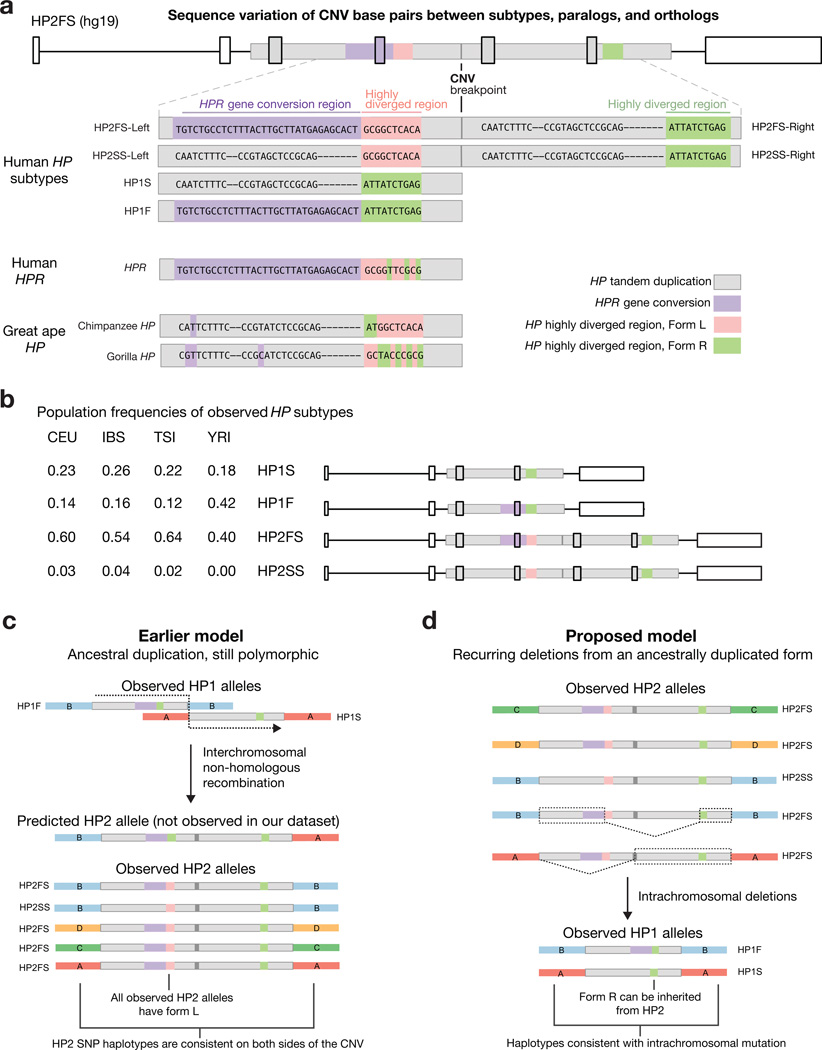
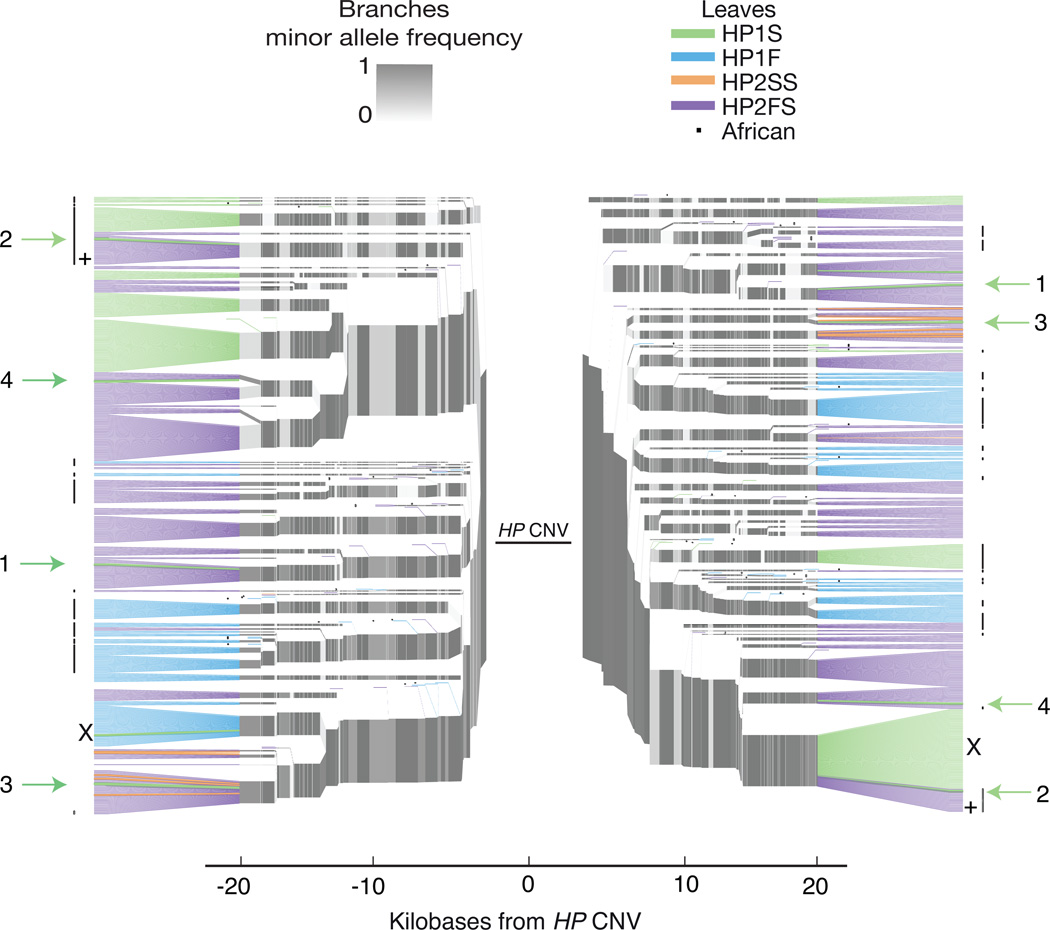
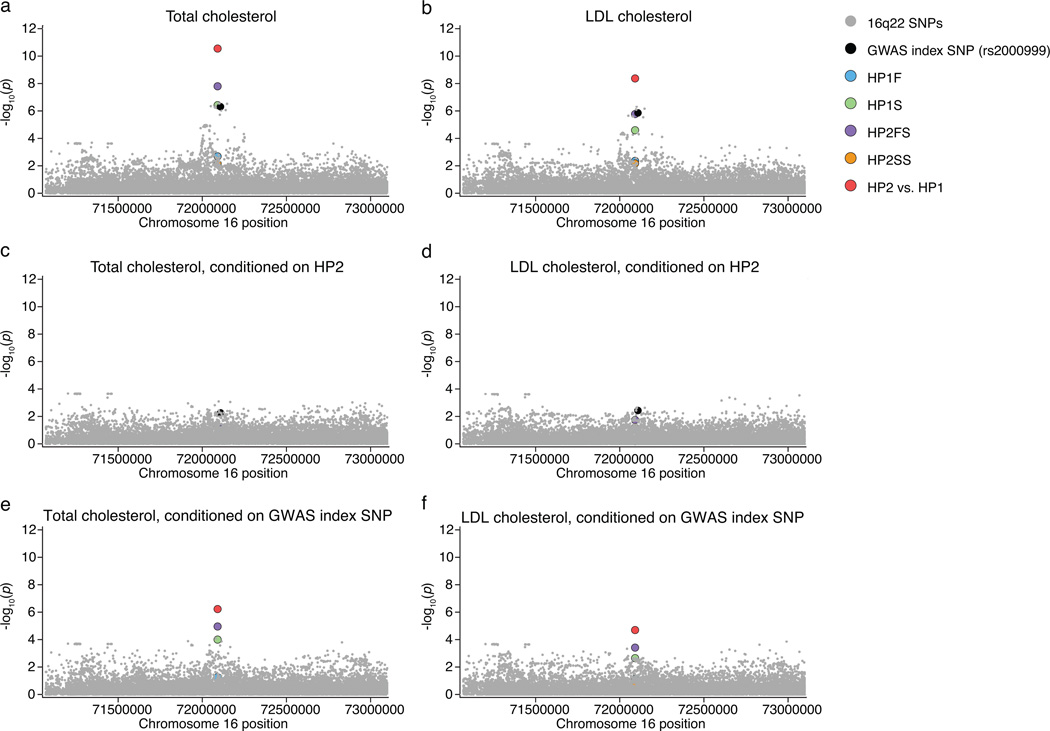
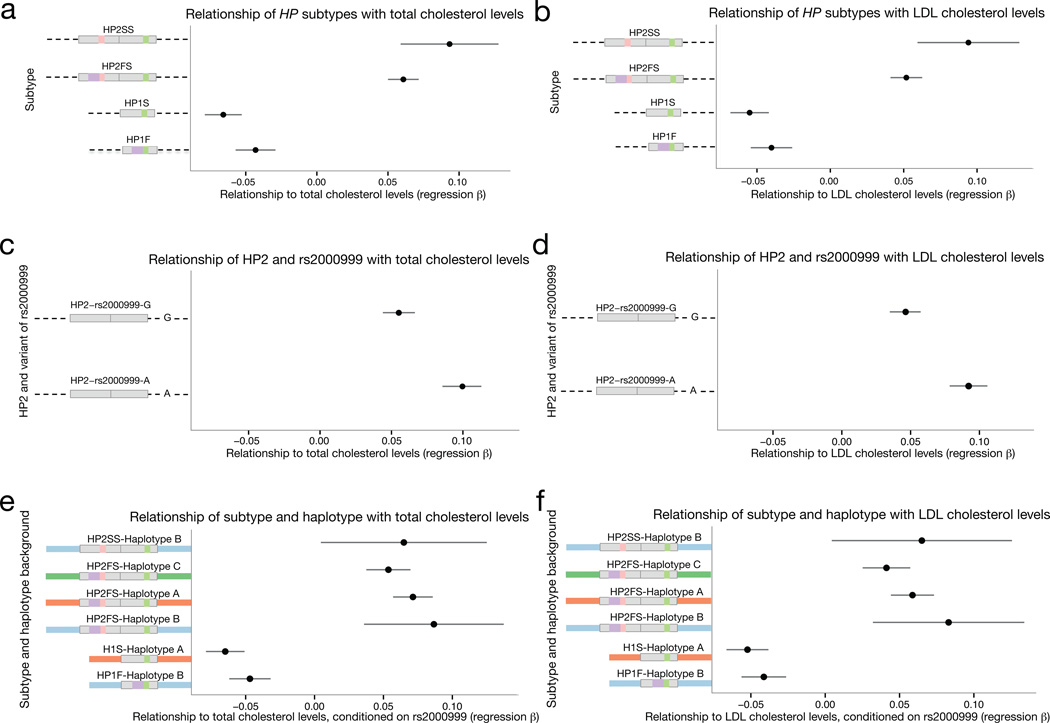
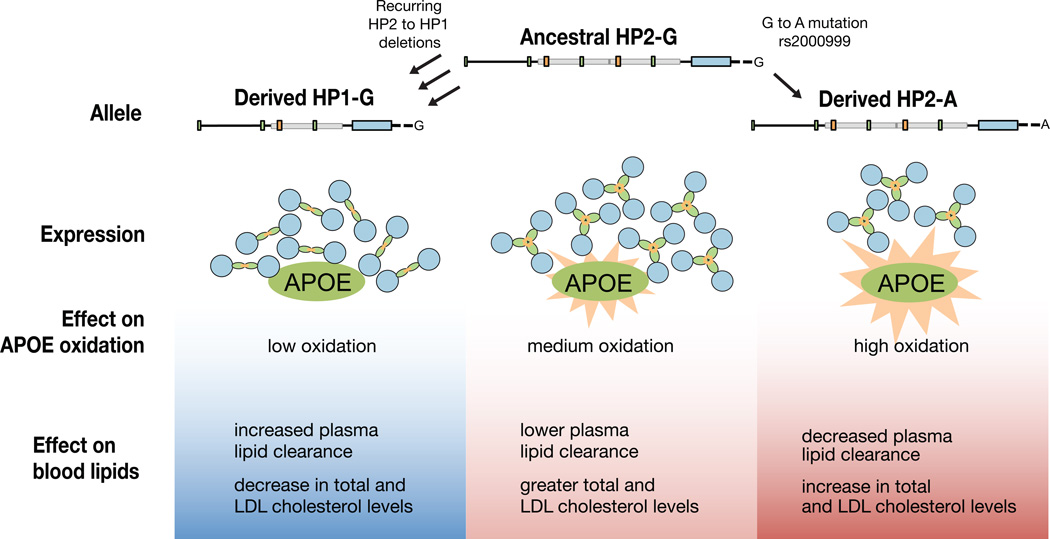
Comment in
-
Recurrent mutation at the classical haptoglobin structural polymorphism.Nat Genet. 2016 Apr;48(4):347-8. doi: 10.1038/ng.3534. Nat Genet. 2016. PMID: 27023774
References
-
- Langlois MR, Delanghe JR. Biological and clinical significance of haptoglobin polymorphism in humans. Clinical Chemistry. 1996;42:1589–1600. - PubMed
-
- Smithies O, Walker NF. Genetic control of some serum proteins in normal humans. Nature. 1955;176:1265–1266. - PubMed
-
- Wejman JC, Hovsepian D, Wall JS, Hainfeld JF, Greer J. Structure and assembly of haptoglobin polymers by electron microscopy. J Mol Biol. 1984;174:343–368. - PubMed
-
- Nielsen MJ, Moestrup SK. Receptor targeting of hemoglobin mediated by the haptoglobins: roles beyond heme scavenging. Blood. 2009;114:764–771. - PubMed
Methods-only references
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
