

Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade
- PMID: 26598659
- PMCID: PMC4702960
- DOI: 10.1073/pnas.1510461112
Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade
Erratum in
-
Correction for Boothby et al., Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade.Proc Natl Acad Sci U S A. 2016 Sep 6;113(36):E5364. doi: 10.1073/pnas.1613046113. Epub 2016 Aug 22. Proc Natl Acad Sci U S A. 2016. PMID: 27551073 Free PMC article. No abstract available.
Abstract
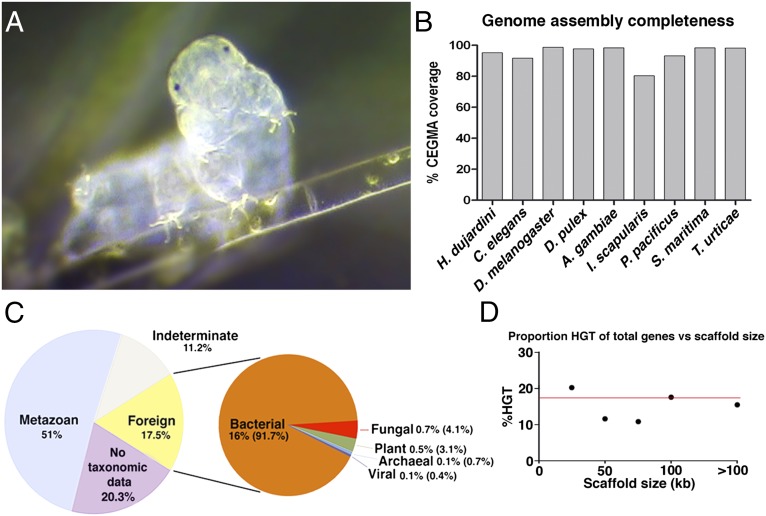
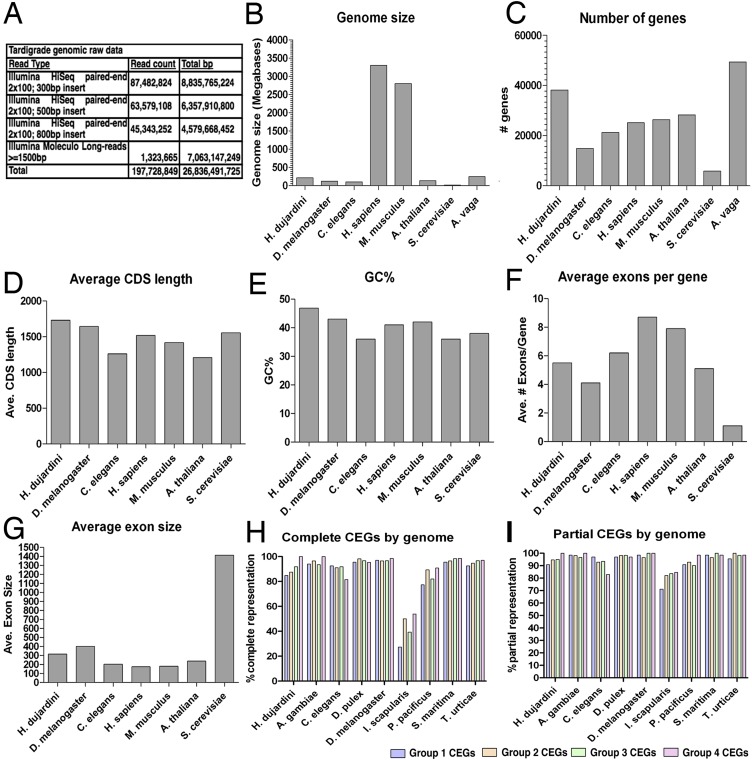
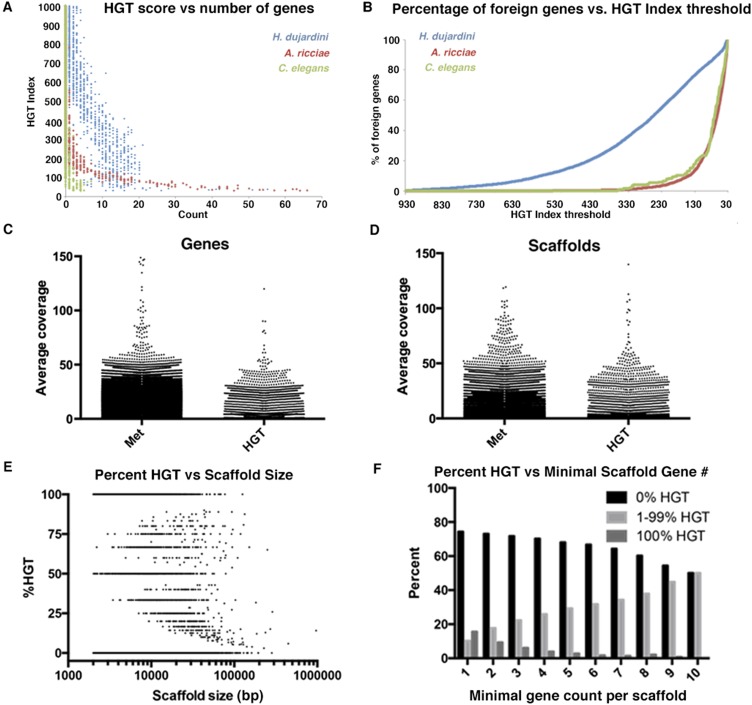
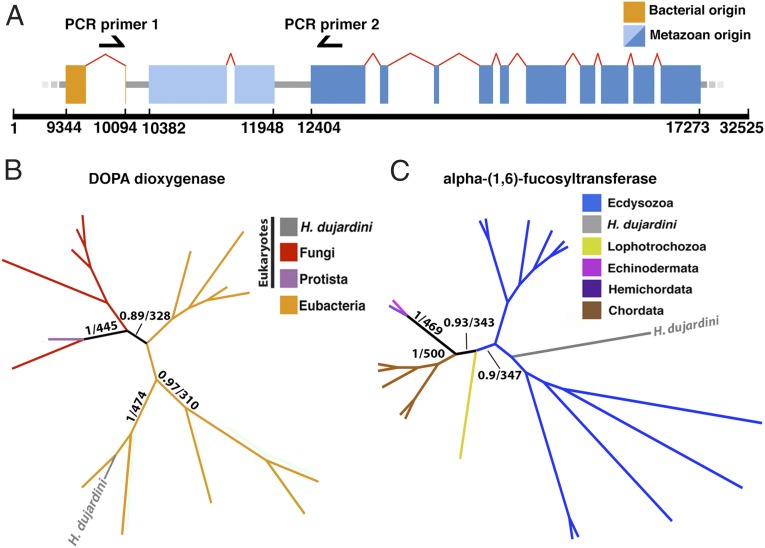
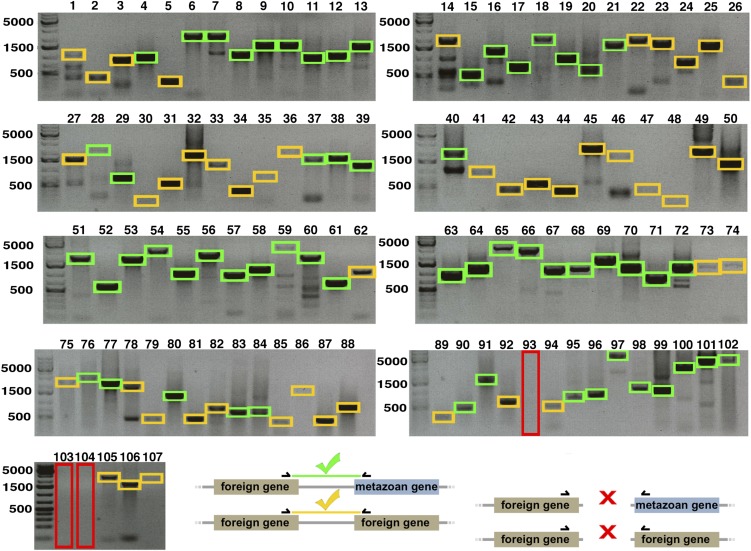
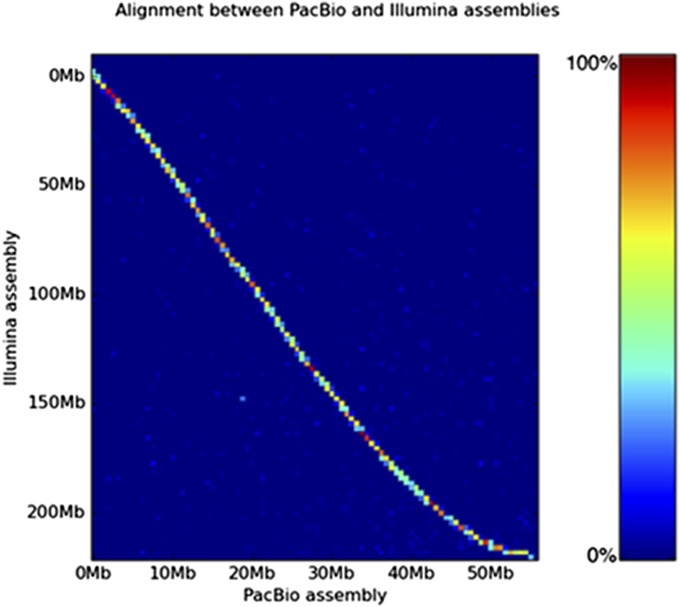
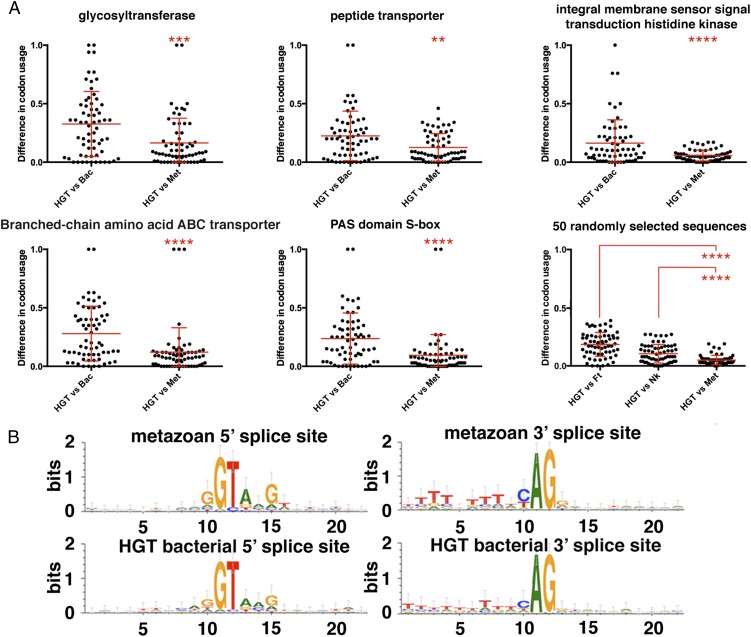
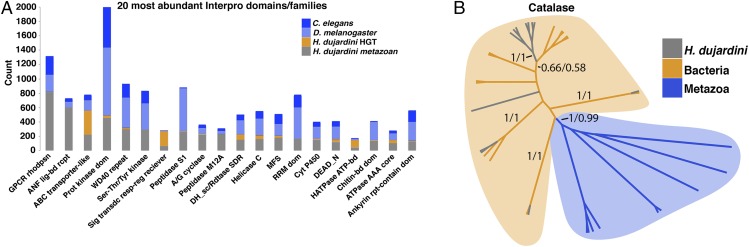
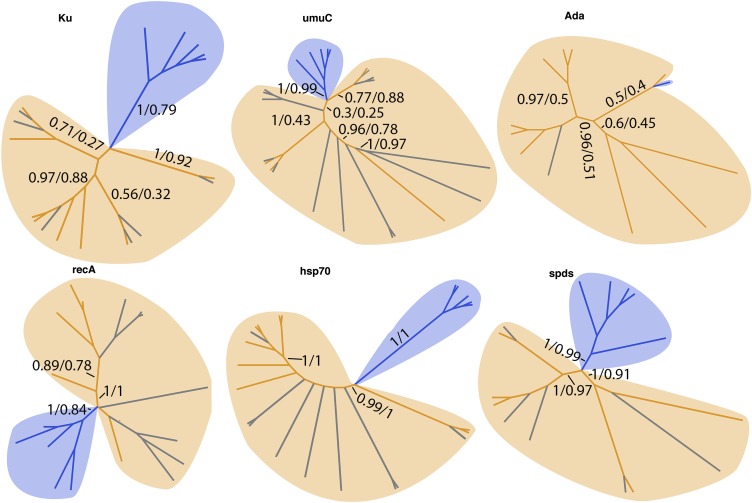
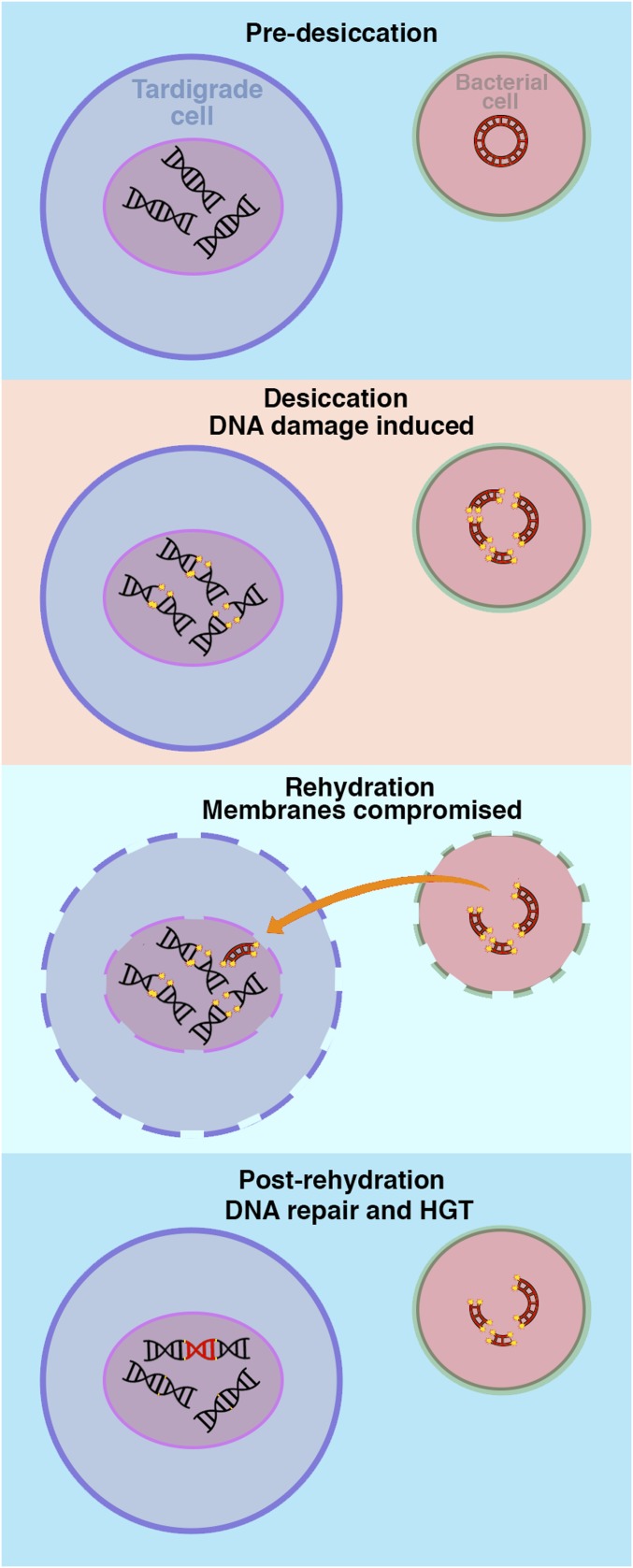
Horizontal gene transfer (HGT), or the transfer of genes between species, has been recognized recently as more pervasive than previously suspected. Here, we report evidence for an unprecedented degree of HGT into an animal genome, based on a draft genome of a tardigrade, Hypsibius dujardini. Tardigrades are microscopic eight-legged animals that are famous for their ability to survive extreme conditions. Genome sequencing, direct confirmation of physical linkage, and phylogenetic analysis revealed that a large fraction of the H. dujardini genome is derived from diverse bacteria as well as plants, fungi, and Archaea. We estimate that approximately one-sixth of tardigrade genes entered by HGT, nearly double the fraction found in the most extreme cases of HGT into animals known to date. Foreign genes have supplemented, expanded, and even replaced some metazoan gene families within the tardigrade genome. Our results demonstrate that an unexpectedly large fraction of an animal genome can be derived from foreign sources. We speculate that animals that can survive extremes may be particularly prone to acquiring foreign genes.
Keywords: genome; horizontal gene transfer; lateral gene transfer; stress tolerance; tardigrade.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Reply to Bemm et al. and Arakawa: Identifying foreign genes in independent Hypsibius dujardini genome assemblies.Proc Natl Acad Sci U S A. 2016 May 31;113(22):E3058-61. doi: 10.1073/pnas.1601149113. Epub 2016 May 12. Proc Natl Acad Sci U S A. 2016. PMID: 27173900 Free PMC article. No abstract available.
-
No evidence for extensive horizontal gene transfer from the draft genome of a tardigrade.Proc Natl Acad Sci U S A. 2016 May 31;113(22):E3057. doi: 10.1073/pnas.1602711113. Epub 2016 May 12. Proc Natl Acad Sci U S A. 2016. PMID: 27173901 Free PMC article. No abstract available.
-
Genome of a tardigrade: Horizontal gene transfer or bacterial contamination?Proc Natl Acad Sci U S A. 2016 May 31;113(22):E3054-6. doi: 10.1073/pnas.1525116113. Epub 2016 May 12. Proc Natl Acad Sci U S A. 2016. PMID: 27173902 Free PMC article. No abstract available.
References
-
- Kinchin IM. The Biology of Tardigrades. Portland Press; London: 1994.
-
- Wright JC. Cryptobiosis 300 years on from van Leuwenhoek: What have we learned about tardigrades? Zool Anz. 2001;240(3-4):563–582.
-
- Becquerel P. La suspension de la vie au dessous de 1/20 K absolu par demagnetization adiabatique de l’alun de fer dans le vide les plus elève. CR Hebd Seances Acad Sci. 1950;231:261–263. French.
-
- Rahm PG. Biologische und physiologische Beiträge zur Kenntnis der Moosfauna. Zeitschrift Allg Physiologie. 1921;20:1–35. German.
-
- Jönsson KI, Harms-Ringdahl M, Torudd J. Radiation tolerance in the eutardigrade Richtersius coronifer. Int J Radiat Biol. 2005;81(9):649–656. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
