

Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics
- PMID: 15378069
- PMCID: PMC4372090
- DOI: 10.1038/nature02976
Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics
Abstract
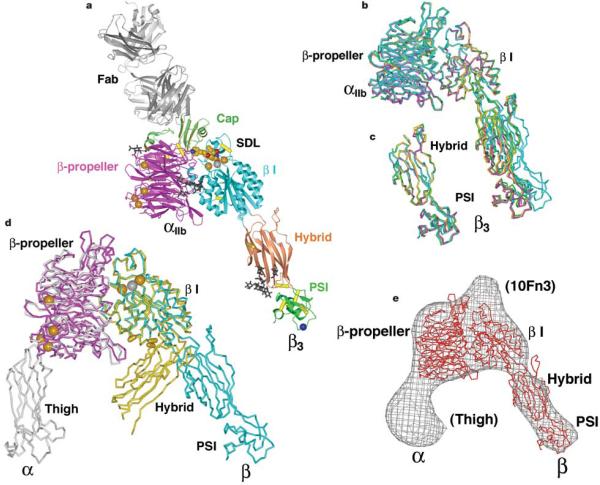
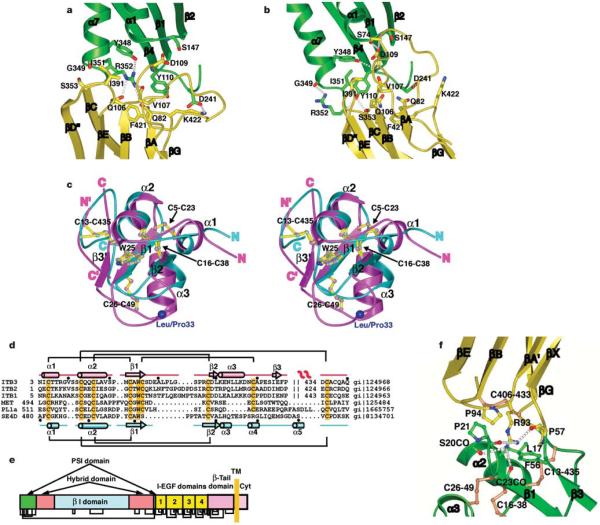
Integrins are important adhesion receptors in all Metazoa that transmit conformational change bidirectionally across the membrane. Integrin alpha and beta subunits form a head and two long legs in the ectodomain and span the membrane. Here, we define with crystal structures the atomic basis for allosteric regulation of the conformation and affinity for ligand of the integrin ectodomain, and how fibrinogen-mimetic therapeutics bind to platelet integrin alpha(IIb)beta3. Allostery in the beta3 I domain alters three metal binding sites, associated loops and alpha1- and alpha7-helices. Piston-like displacement of the alpha7-helix causes a 62 degrees reorientation between the beta3 I and hybrid domains. Transmission through the rigidly connected plexin/semaphorin/integrin (PSI) domain in the upper beta3 leg causes a 70 A separation between the knees of the alpha and beta legs. Allostery in the head thus disrupts interaction between the legs in a previously described low-affinity bent integrin conformation, and leg extension positions the high-affinity head far above the cell surface.
Figures



Comment in
-
Cell biology: adhesion articulated.Nature. 2004 Nov 4;432(7013):27-8. doi: 10.1038/432027a. Nature. 2004. PMID: 15525967 No abstract available.
References
-
- Hughes PE, Pfaff M. Integrin affinity modulation. Trends Cell Biol. 1998;8:359–364. - PubMed
-
- Takagi J, Springer TA. Integrin activation and structural rearrangement. Immunol. Rev. 2002;186:141–163. - PubMed
-
- Takagi J, Petre BM, Walz T, Springer TA. Global conformational rearrangements in integrin extracellular domains in outside-in and inside-out signaling. Cell. 2002;110:599–611. - PubMed
-
- Springer TA, Wang J.-h. In: Cell Surface Receptors. Garcia KC), editor. Elsevier; San Diego: 2004.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases