

Evolution of function in the "two dinucleotide binding domains" flavoproteins
- PMID: 17658942
- PMCID: PMC1924876
- DOI: 10.1371/journal.pcbi.0030121
Evolution of function in the "two dinucleotide binding domains" flavoproteins
Abstract
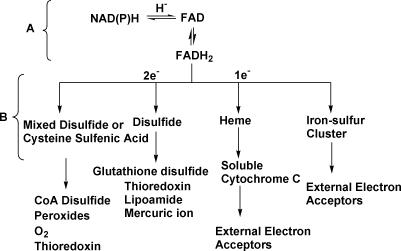
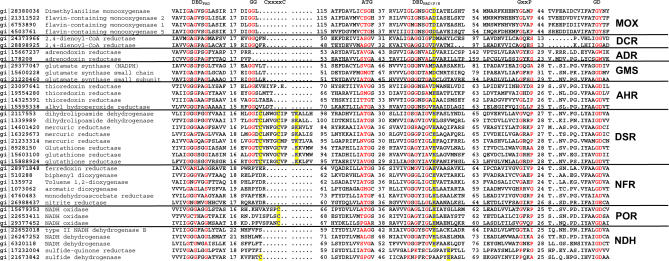
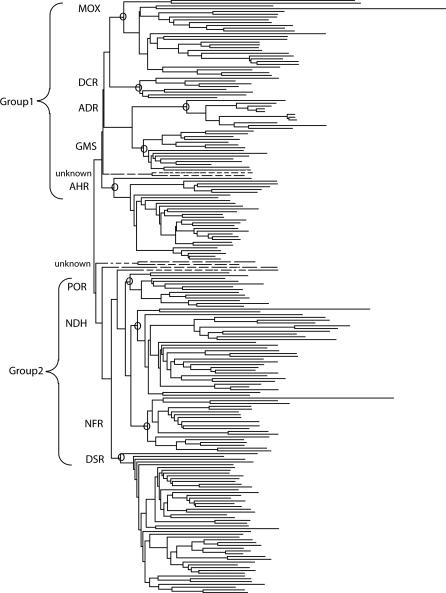
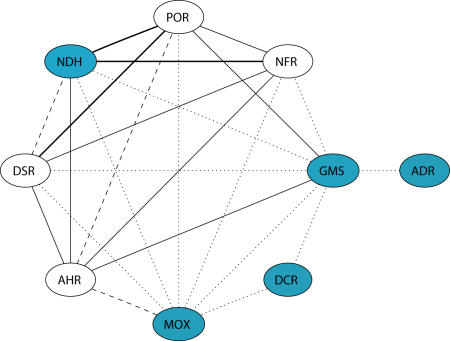
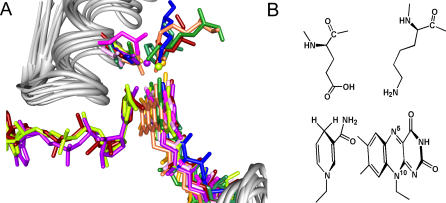
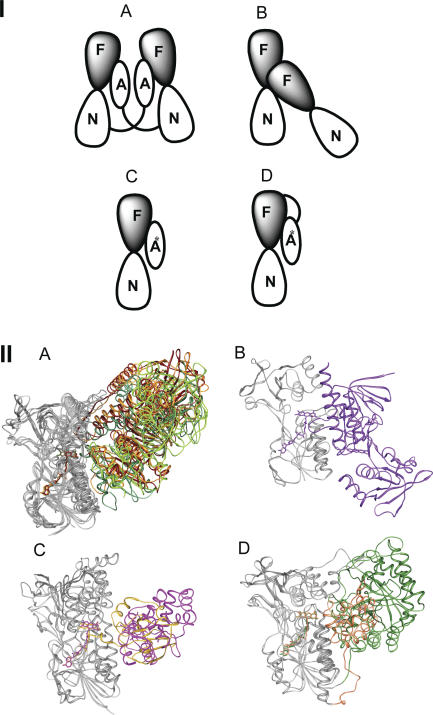
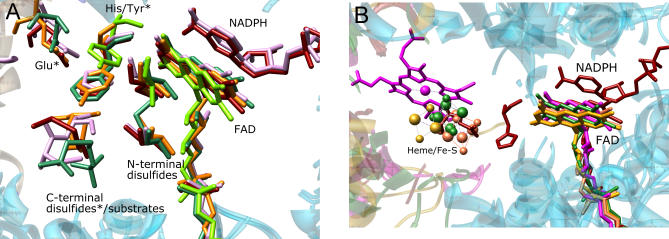
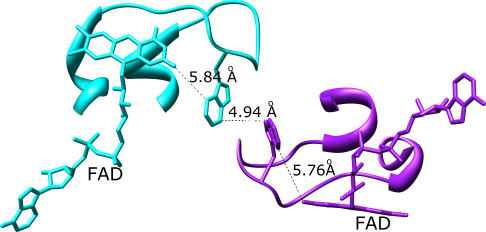
Structural and biochemical constraints force some segments of proteins to evolve more slowly than others, often allowing identification of conserved structural or sequence motifs that can be associated with substrate binding properties, chemical mechanisms, and molecular functions. We have assessed the functional and structural constraints imposed by cofactors on the evolution of new functions in a superfamily of flavoproteins characterized by two-dinucleotide binding domains, the "two dinucleotide binding domains" flavoproteins (tDBDF) superfamily. Although these enzymes catalyze many different types of oxidation/reduction reactions, each is initiated by a stereospecific hydride transfer reaction between two cofactors, a pyridine nucleotide and flavin adenine dinucleotide (FAD). Sequence and structural analysis of more than 1,600 members of the superfamily reveals new members and identifies details of the evolutionary connections among them. Our analysis shows that in all of the highly divergent families within the superfamily, these cofactors adopt a conserved configuration optimal for stereospecific hydride transfer that is stabilized by specific interactions with amino acids from several motifs distributed among both dinucleotide binding domains. The conservation of cofactor configuration in the active site restricts the pyridine nucleotide to interact with FAD from the re-side, limiting the flow of electrons from the re-side to the si-side. This directionality of electron flow constrains interactions with the different partner proteins of different families to occur on the same face of the cofactor binding domains. As a result, superimposing the structures of tDBDFs aligns not only these interacting proteins, but also their constituent electron acceptors, including heme and iron-sulfur clusters. Thus, not only are specific aspects of the cofactor-directed chemical mechanism conserved across the superfamily, the constraints they impose are manifested in the mode of protein-protein interactions. Overlaid on this foundation of conserved interactions, nature has conscripted different protein partners to serve as electron acceptors, thereby generating diversification of function across the superfamily.
Conflict of interest statement
Figures
References
-
- May AC, Johnson MS, Rufino SD, Wako H, Zhu ZY, et al. The recognition of protein structure and function from sequence: Adding value to genome data. Philos Trans R Soc Lond B Biol Sci. 1994;344:373–381. - PubMed
-
- Babbitt PC, Gerlt JA. Understanding enzyme superfamilies. Chemistry as the fundamental determinant in the evolution of new catalytic activities. J Biol Chem. 1997;272:30591–30594. - PubMed
-
- Copley SD, Novak WR, Babbitt PC. Divergence of function in the thioredoxin fold suprafamily: Evidence for evolution of peroxiredoxins from a thioredoxin-like ancestor. Biochemistry. 2004;43:13981–13995. - PubMed
-
- Seibert CM, Raushel FM. Structural and catalytic diversity within the amidohydrolase superfamily. Biochemistry. 2005;44:6383–6391. - PubMed
-
- Burroughs AM, Allen KN, Dunaway-Mariano D, Aravind L. Evolutionary genomics of the HAD superfamily: Understanding the structural adaptations and catalytic diversity in a superfamily of phosphoesterases and allied enzymes. J Mol Biol. 2006;361:1003–1034. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
