

Crystal structures of a template-independent DNA polymerase: murine terminal deoxynucleotidyltransferase
- PMID: 11823435
- PMCID: PMC125842
- DOI: 10.1093/emboj/21.3.427
Crystal structures of a template-independent DNA polymerase: murine terminal deoxynucleotidyltransferase
Abstract
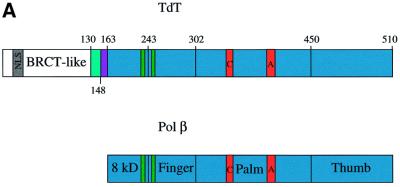
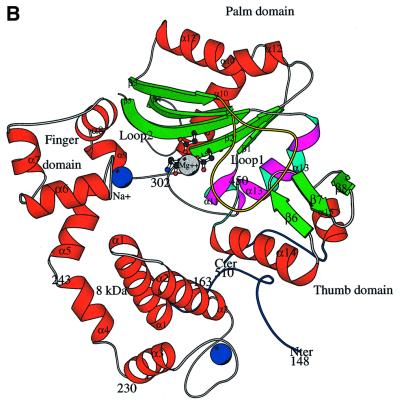
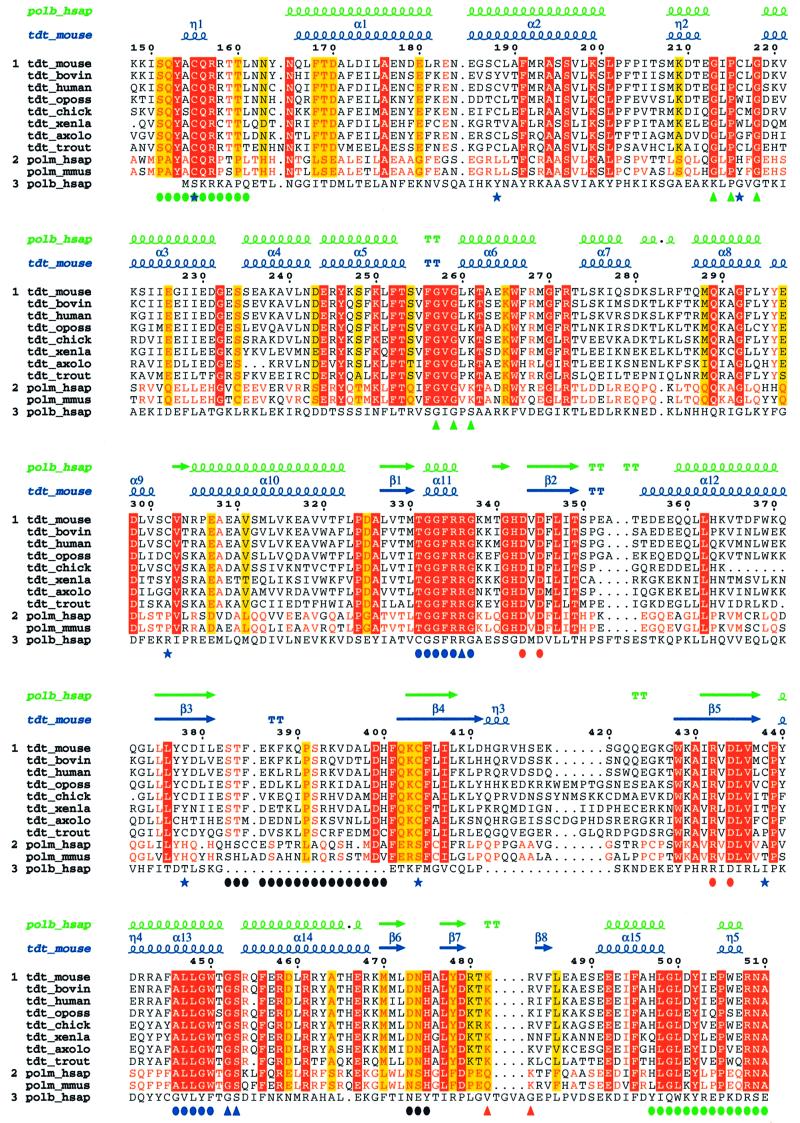
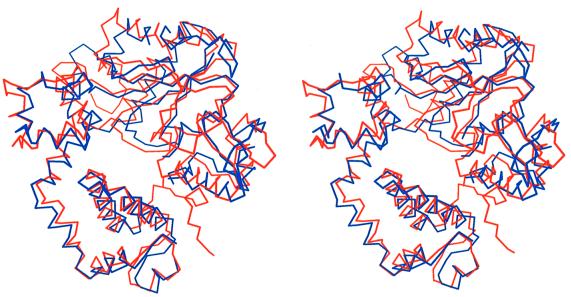
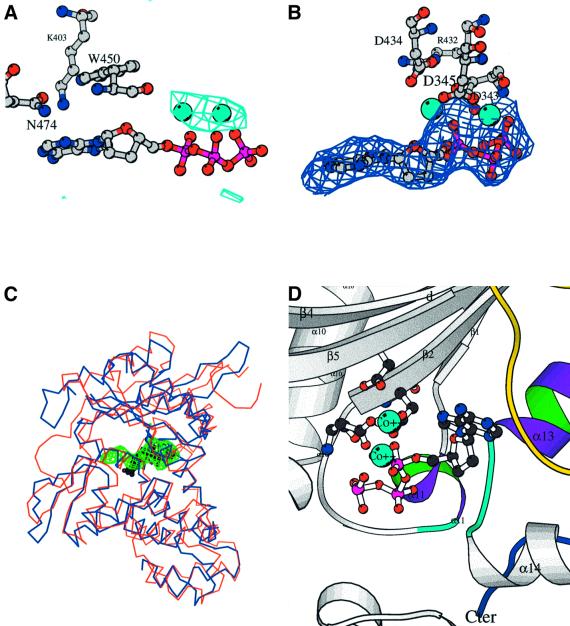
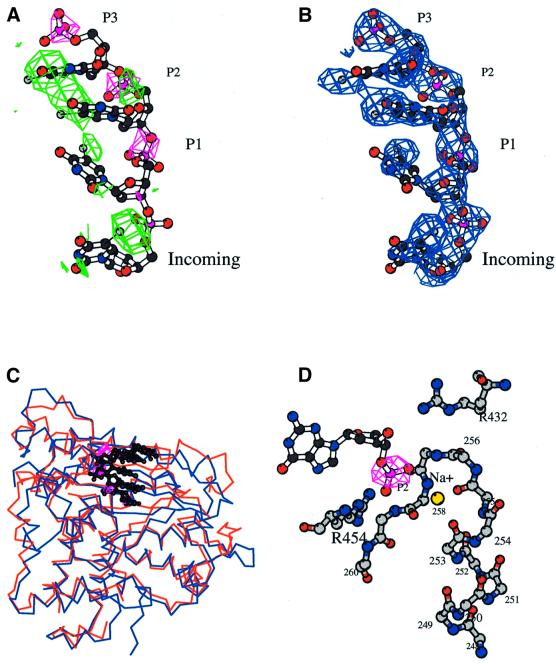
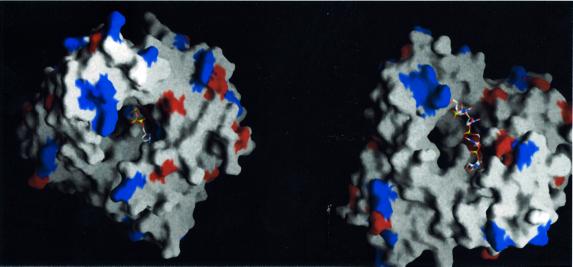
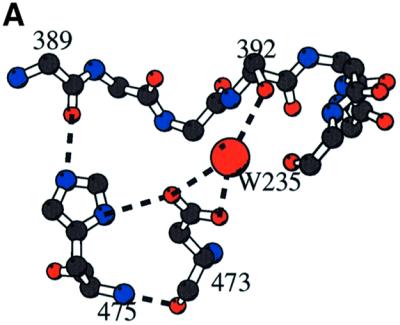
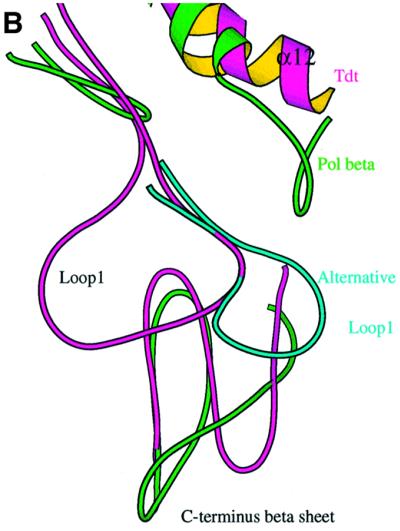
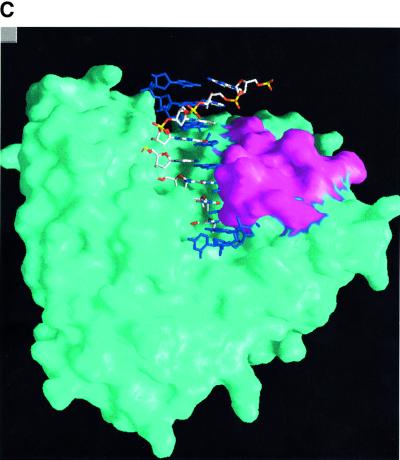
The crystal structure of the catalytic core of murine terminal deoxynucleotidyltransferase (TdT) at 2.35 A resolution reveals a typical DNA polymerase beta-like fold locked in a closed form. In addition, the structures of two different binary complexes, one with an oligonucleotide primer and the other with an incoming ddATP-Co(2+) complex, show that the substrates and the two divalent ions in the catalytic site are positioned in TdT in a manner similar to that described for the human DNA polymerase beta ternary complex, suggesting a common two metal ions mechanism of nucleotidyl transfer in these two proteins. The inability of TdT to accommodate a template strand can be explained by steric hindrance at the catalytic site caused by a long lariat-like loop, which is absent in DNA polymerase beta. However, displacement of this discriminating loop would be sufficient to unmask a number of evolutionarily conserved residues, which could then interact with a template DNA strand. The present structure can be used to model the recently discovered human polymerase mu, with which it shares 43% sequence identity.
Figures
References
-
- Bard J., Zhelovsky,A.M., Helmling,S., Earnest,T.N., Moore,C.L. and Bohm,A. (2000) Structure of yeast poly(A) polymerase alone and in complex with 3′dATP. Science, 289, 1346–1349. - PubMed
-
- Beard W.A. and Wilson,S.H. (1998) Structural insights into DNA polymerase β fidelity: hold it tight if you want it right. Chem. Biol., 5, R7–R13. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
