

Crystal structures of a pantothenate synthetase from M. tuberculosis and its complexes with substrates and a reaction intermediate
- PMID: 12717031
- PMCID: PMC2323879
- DOI: 10.1110/ps.0241803
Crystal structures of a pantothenate synthetase from M. tuberculosis and its complexes with substrates and a reaction intermediate
Abstract
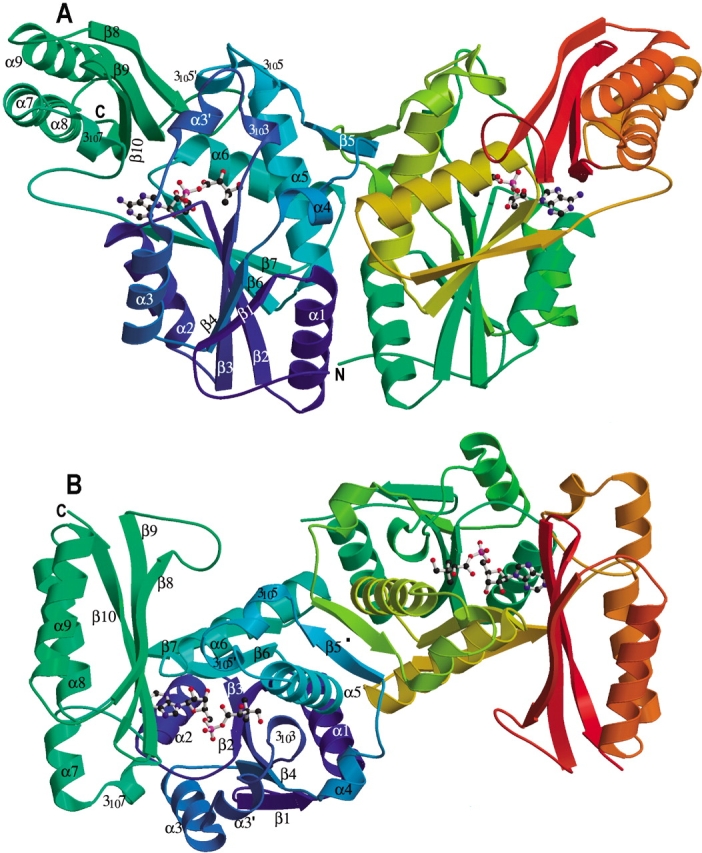
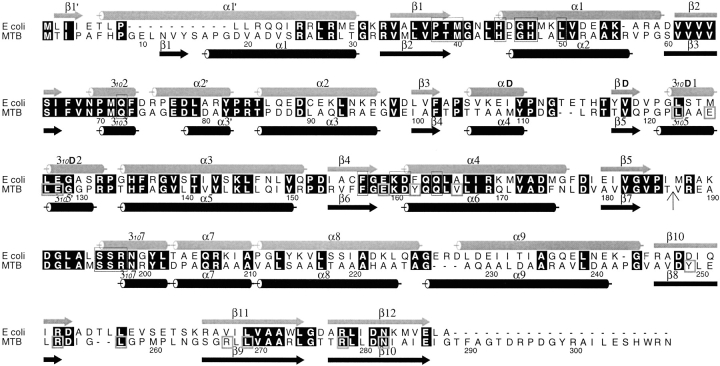
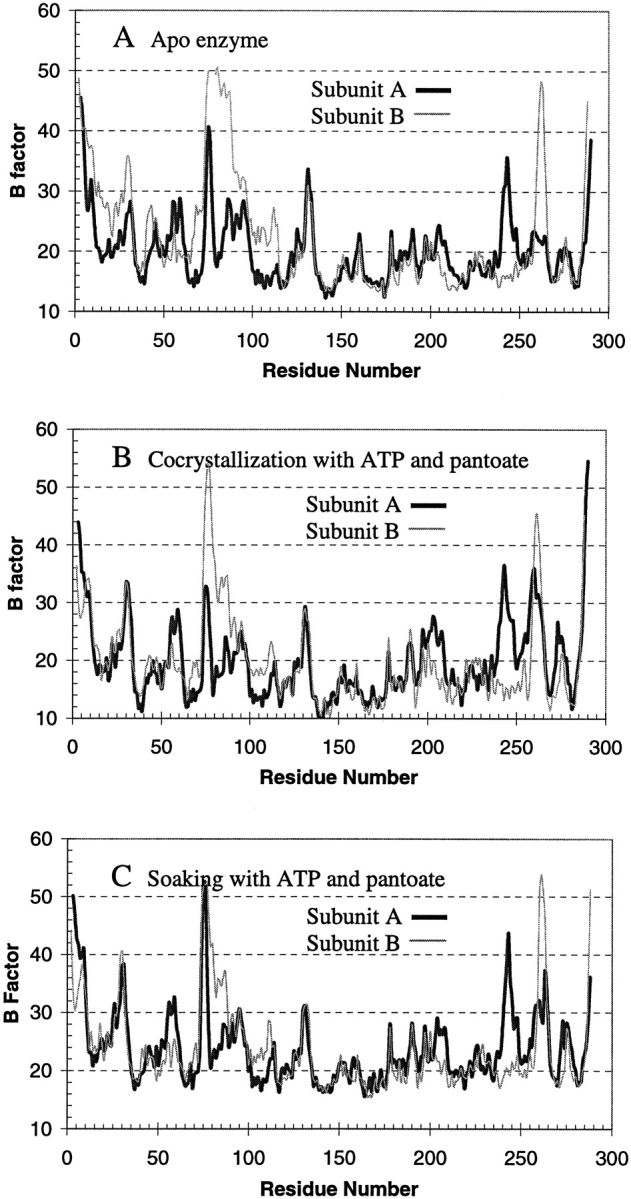
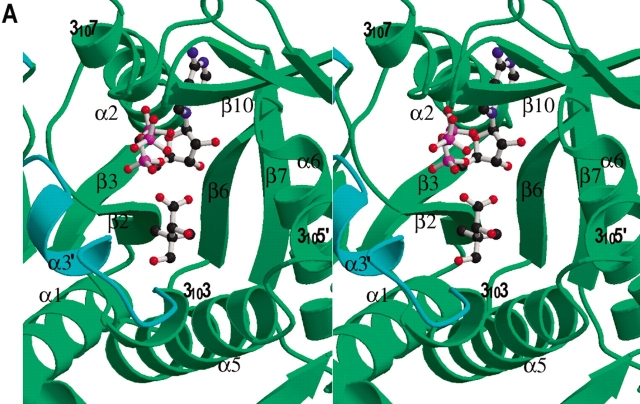
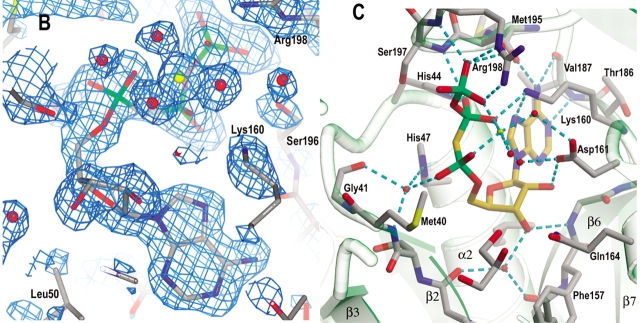
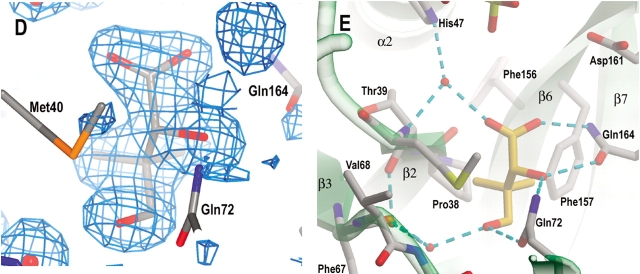
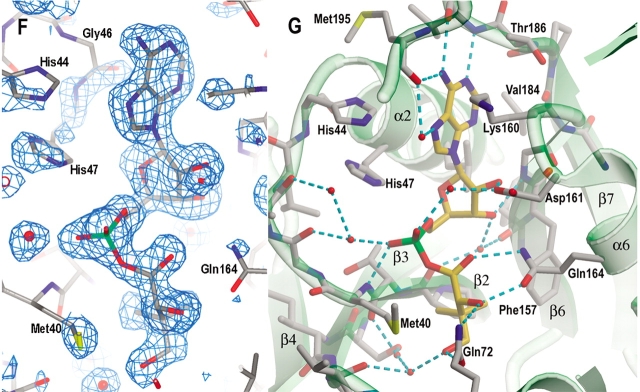
Pantothenate biosynthesis is essential for the virulence of Mycobacterium tuberculosis, and this pathway thus presents potential drug targets against tuberculosis. We determined the crystal structure of pantothenate synthetase (PS) from M. tuberculosis, and its complexes with AMPCPP, pantoate, and a reaction intermediate, pantoyl adenylate, with resolutions from 1.6 to 2 A. PS catalyzes the ATP-dependent condensation of pantoate and beta-alanine to form pantothenate. Its structure reveals a dimer, and each subunit has two domains with tight association between domains. The active-site cavity is on the N-terminal domain, partially covered by the C-terminal domain. One wall of the active site cavity is flexible, which allows the bulky AMPCPP to diffuse into the active site to nearly full occupancy when crystals are soaked in solutions containing AMPCPP. Crystal structures of the complexes with AMPCPP and pantoate indicate that the enzyme binds ATP and pantoate tightly in the active site, and brings the carboxyl oxygen of pantoate near the alpha-phosphorus atom of ATP for an in-line nucleophilic attack. When crystals were soaked with, or grown in the presence of, both ATP and pantoate, a reaction intermediate, pantoyl adenylate, is found in the active site. The flexible wall of the active site cavity becomes ordered when the intermediate is in the active site, thus protecting it from being hydrolyzed. Binding of beta-alanine can occur only after pantoyl adenylate is formed inside the active site cavity. The tight binding of the intermediate pantoyl adenylate suggests that nonreactive analogs of pantoyl adenylate may be inhibitors of the PS enzyme with high affinity and specificity.
Figures
References
-
- Barker, D.G. and Winter, G. 1982. Conserved cysteine and histidine residues in the structures of the tyrosyl and methionyl-tRNA synthetases. FEBS Lett. 145 191–193. - PubMed
-
- Barton, G.J. 1993. ALSCRIPT: A tool to format multiple sequence alignments. Protein Eng. 6 37–40. - PubMed
-
- Brunger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P., Grosse-Kunstleve, R.W., Jiang, J.-S., Kuszewski, J., Nilges, N., Pannu, N.S., et al. 1998. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D54 905–921. - PubMed
-
- CCP4 (Collaborative Computational Project Number 4). 1994. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D50 760–763. - PubMed
-
- Cowtan, K. 1994. DM: An automated procedure for phase improvement by density modification. Joint CCP4 ESF-EACBM Newslett. Protein Crystallogr. 31 34–38.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
