

Nanosensor detection of an immunoregulatory tryptophan influx/kynurenine efflux cycle
- PMID: 17896864
- PMCID: PMC1988858
- DOI: 10.1371/journal.pbio.0050257
Nanosensor detection of an immunoregulatory tryptophan influx/kynurenine efflux cycle
Abstract
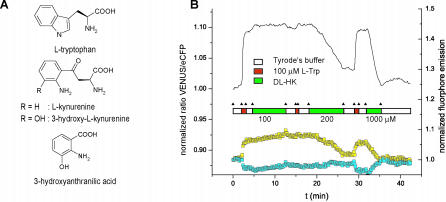
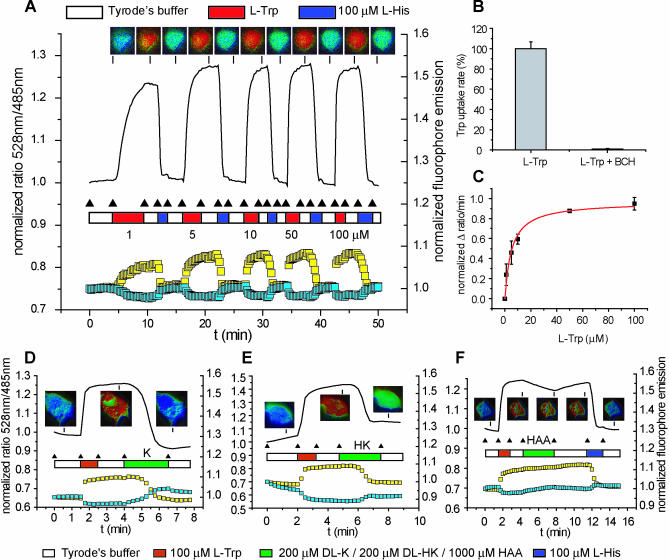
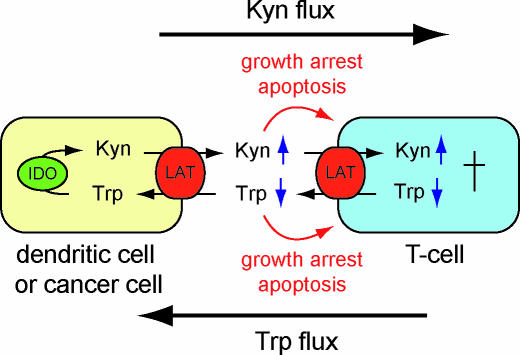
Mammalian cells rely on cellular uptake of the essential amino acid tryptophan. Tryptophan sequestration by up-regulation of the key enzyme for tryptophan degradation, indoleamine 2,3-dioxygenase (IDO), e.g., in cancer and inflammation, is thought to suppress the immune response via T cell starvation. Additionally, the excreted tryptophan catabolites (kynurenines) induce apoptosis of lymphocytes. Whereas tryptophan transport systems have been identified, the molecular nature of kynurenine export remains unknown. To measure cytosolic tryptophan steady-state levels and flux in real time, we developed genetically encoded fluorescence resonance energy transfer nanosensors (FLIPW). The transport properties detected by FLIPW in KB cells, a human oral cancer cell line, and COS-7 cells implicate LAT1, a transporter that is present in proliferative tissues like cancer, in tryptophan uptake. Importantly, we found that this transport system mediates tryptophan/kynurenine exchange. The tryptophan influx/kynurenine efflux cycle couples tryptophan starvation to elevation of kynurenine serum levels, providing a two-pronged induction of apoptosis in neighboring cells. The strict coupling protects cells that overproduce IDO from kynurenine accumulation. Consequently, this mechanism may contribute to immunosuppression involved in autoimmunity and tumor immune escape.
Conflict of interest statement
Figures
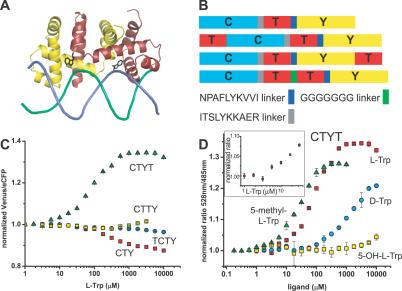
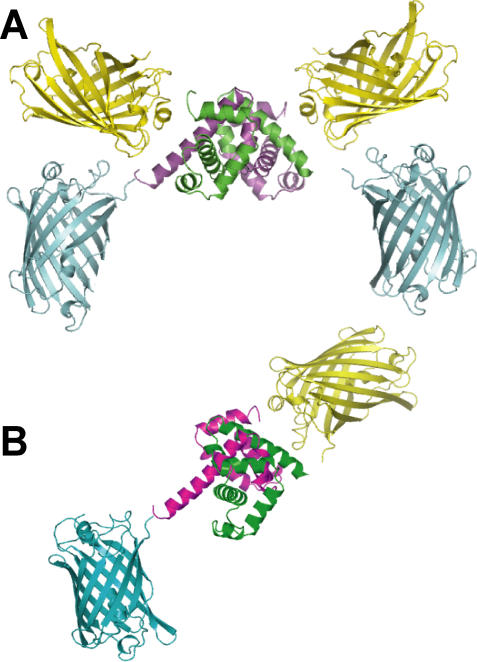
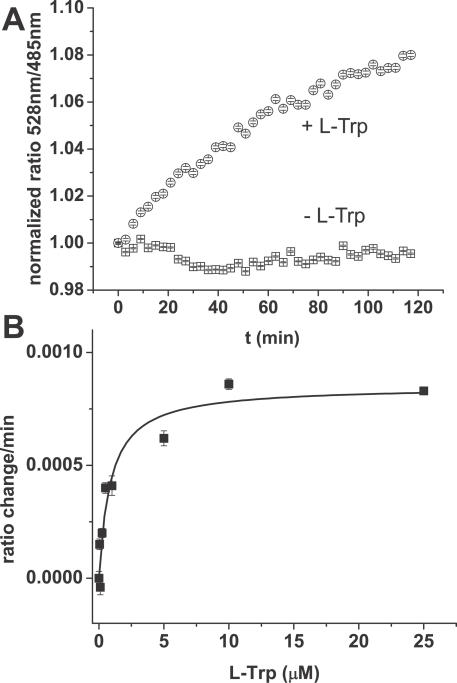
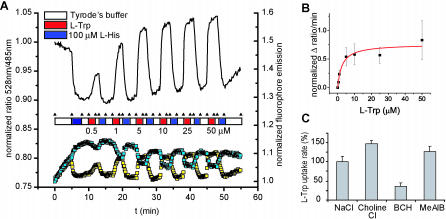
References
-
- Broer S, Cavanaugh JA, Rasko JEJ. Neutral amino acid transport in epithelial cells and its malfunction in Hartnup disorder. Biochem Soc Trans. 2005;33:233–236. - PubMed
-
- Feliubadalo L, Font M, Purroy J, Rousaud F, Estivill X, et al. Non-type I cystinuria caused by mutations in SLC7A9, encoding a subunit (bo,+AT) of rBAT. Nat Genet. 1999;23:52–57. - PubMed
-
- Kim DK, Kanai Y, Matsuo H, Kim JY, Chairoungdua A, et al. The human T-type amino acid transporter-1: Characterization, gene organization, and chromosomal location. Genomics. 2002;79:95–103. - PubMed
-
- Torrents D, Estevez R, Pineda M, Fernandez E, Lloberas J, et al. Identification and characterization of a membrane protein (y+L amino acid transporter-1) that associates with 4F2hc to encode the amino acid transport activity y+L. A candidate gene for lysinuric protein intolerance. J Biol Chem. 1998;273:32437–32445. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
