

Family-specific Kinesin Structures Reveal Neck-linker Length Based on Initiation of the Coiled-coil
- PMID: 27462072
- PMCID: PMC5034036
- DOI: 10.1074/jbc.M116.737577
Family-specific Kinesin Structures Reveal Neck-linker Length Based on Initiation of the Coiled-coil
Abstract
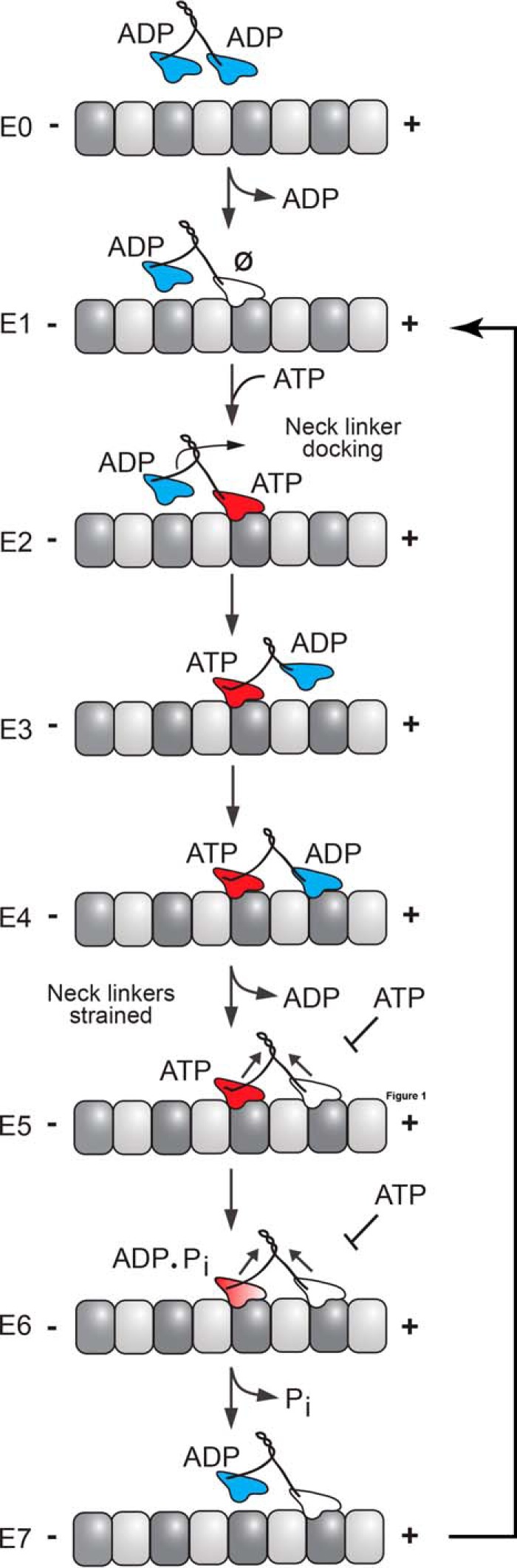
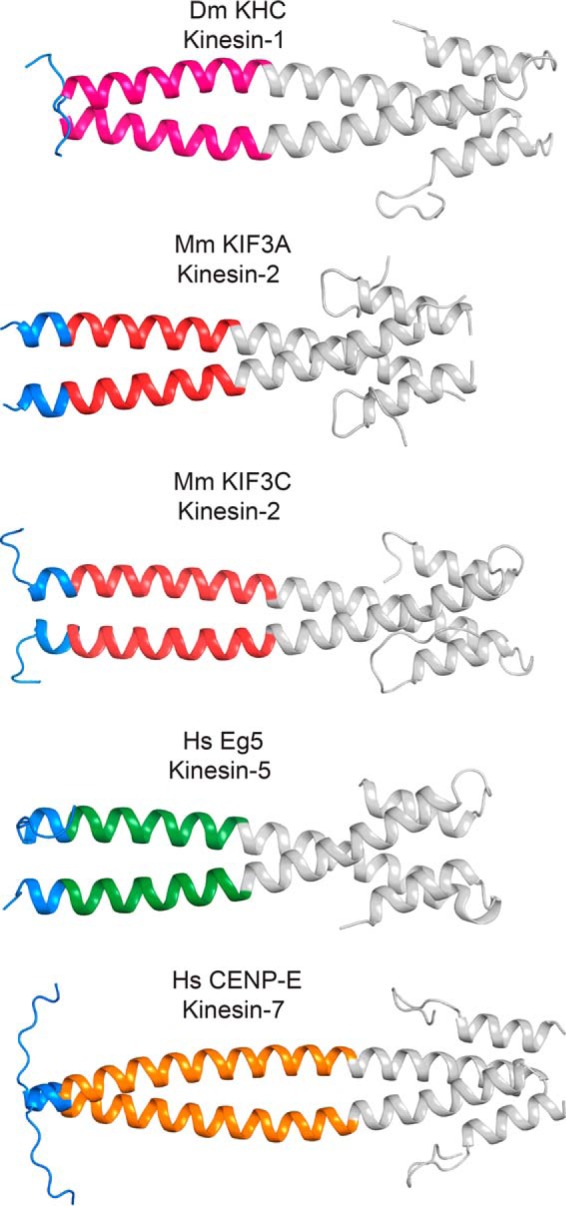
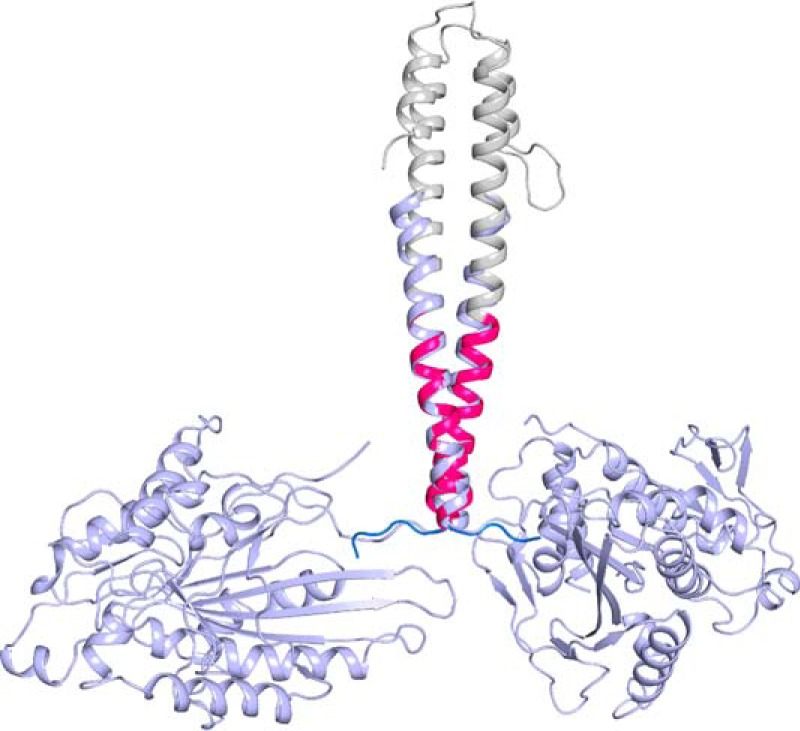
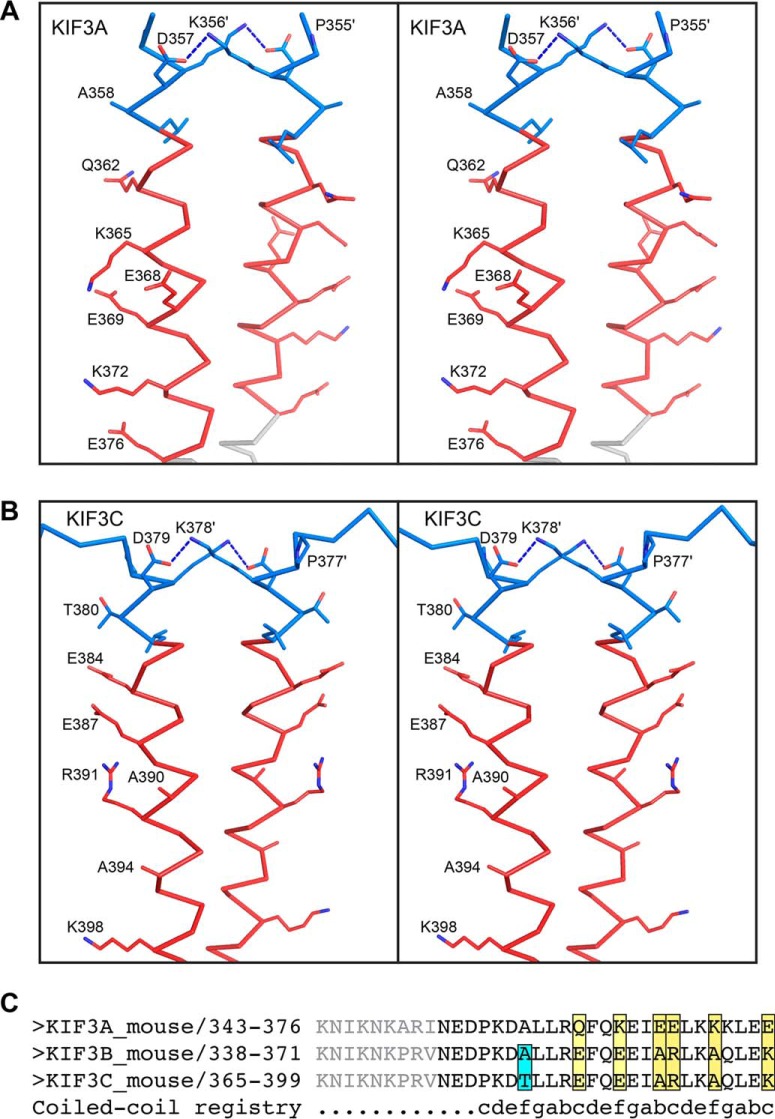
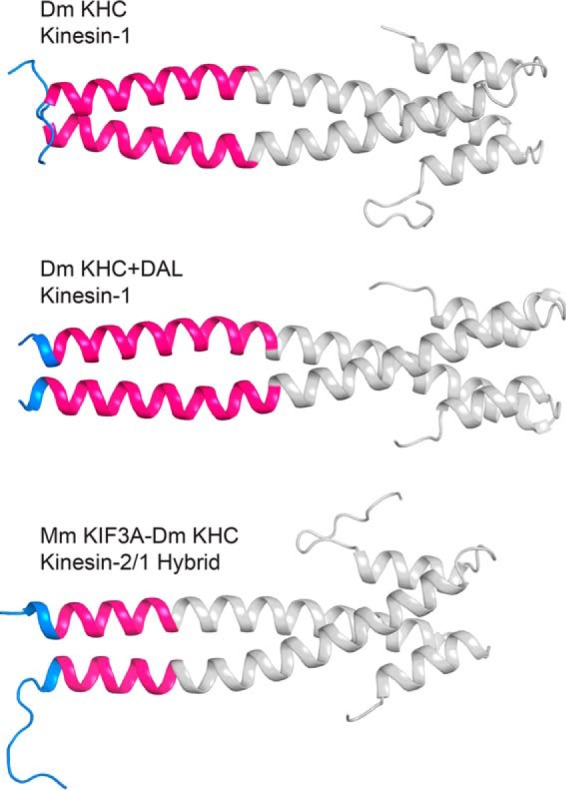
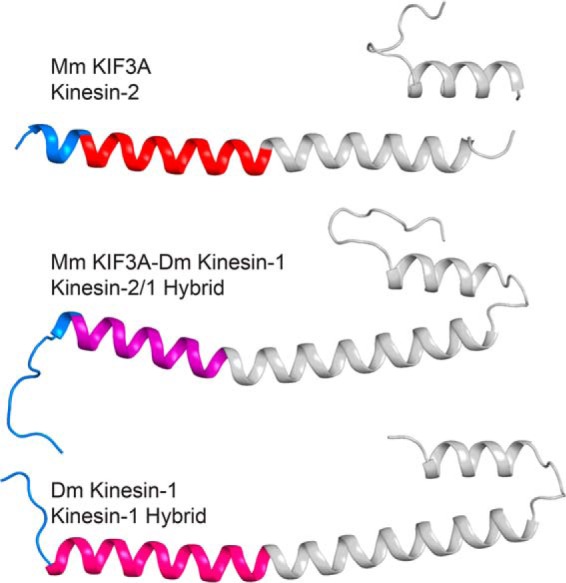
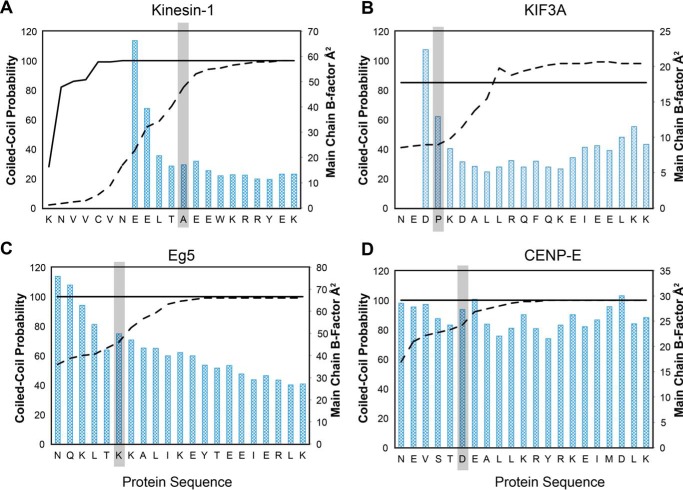
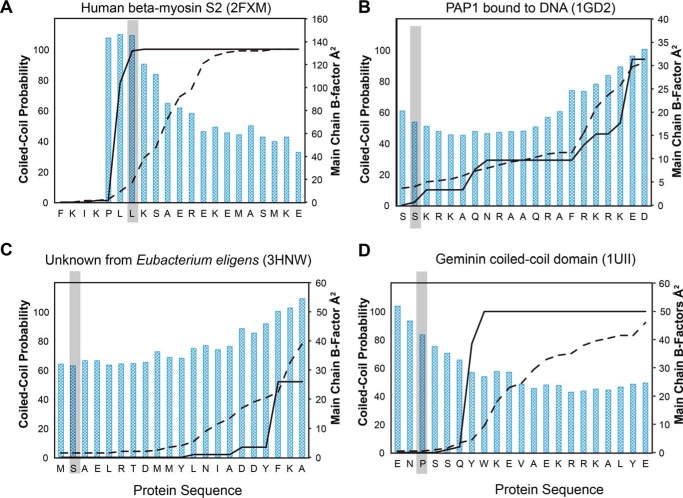
Kinesin-1, -2, -5, and -7 generate processive hand-over-hand 8-nm steps to transport intracellular cargoes toward the microtubule plus end. This processive motility requires gating mechanisms to coordinate the mechanochemical cycles of the two motor heads to sustain the processive run. A key structural element believed to regulate the degree of processivity is the neck-linker, a short peptide of 12-18 residues, which connects the motor domain to its coiled-coil stalk. Although a shorter neck-linker has been correlated with longer run lengths, the structural data to support this hypothesis have been lacking. To test this hypothesis, seven kinesin structures were determined by x-ray crystallography. Each included the neck-linker motif, followed by helix α7 that constitutes the start of the coiled-coil stalk. In the majority of the structures, the neck-linker length differed from predictions because helix α7, which initiates the coiled-coil, started earlier in the sequence than predicted. A further examination of structures in the Protein Data Bank reveals that there is a great disparity between the predicted and observed starting residues. This suggests that an accurate prediction of the start of a coiled-coil is currently difficult to achieve. These results are significant because they now exclude simple comparisons between members of the kinesin superfamily and add a further layer of complexity when interpreting the results of mutagenesis or protein fusion. They also re-emphasize the need to consider factors beyond the kinesin neck-linker motif when attempting to understand how inter-head communication is tuned to achieve the degree of processivity required for cellular function.
Keywords: coiled-coil; kinesin; kinesin neck-linker; microtubule; molecular motor; protein structure; x-ray crystallography.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Crick F. H. (1953) The packing of α-helices: simple coiled-coils. Acta Crystallogr. 6, 689–697
-
- Rackham O. J., Madera M., Armstrong C. T., Vincent T. L., Woolfson D. N., and Gough J. (2010) The evolution and structure prediction of coiled coils across all genomes. J. Mol. Biol. 403, 480–493 - PubMed
-
- Delorenzi M., and Speed T. (2002) An HMM model for coiled-coil domains and a comparison with PSSM-based predictions. Bioinformatics 18, 617–625 - PubMed
-
- Lupas A., Van Dyke M., and Stock J. (1991) Predicting coiled coils from protein sequences. Science 252, 1162–1164 - PubMed
-
- Parry D. A., Fraser R. D., and Squire J. M. (2008) Fifty years of coiled-coils and α-helical bundles: a close relationship between sequence and structure. J. Struct. Biol. 163, 258–269 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
