

NusA interaction with the α subunit of E. coli RNA polymerase is via the UP element site and releases autoinhibition
- PMID: 21742261
- PMCID: PMC3134791
- DOI: 10.1016/j.str.2011.03.024
NusA interaction with the α subunit of E. coli RNA polymerase is via the UP element site and releases autoinhibition
Abstract
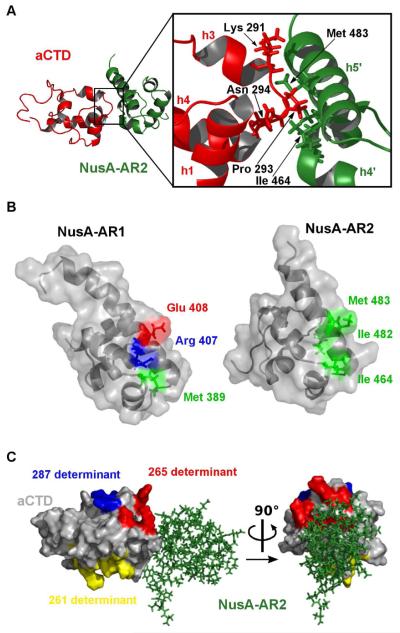
Elongating Escherichia coli RNAP is modulated by NusA protein. The C-terminal domain (CTD) of the RNAP α subunit (αCTD) interacts with the acidic CTD 2 (AR2) of NusA, releasing the autoinhibitory blockade of the NusA S1-KH1-KH2 motif and allowing NusA to bind nascent nut spacer RNA. We determined the solution conformation of the AR2:αCTD complex. The αCTD residues that interface with AR2 are identical to those that recognize UP promoter elements A nusA-ΔAR2 mutation does not affect UP-dependent rrnH transcription initiation in vivo. Instead, the mutation inhibits Rho-dependent transcription termination at phage λtR1, which lies adjacent to the λnutR sequence. The Rho-dependent λtimm terminator, which is not preceded by a λnut sequence, is fully functional. We propose that constitutive binding of NusA-ΔAR2 to λnutR occludes Rho. In addition, the mutation confers a dominant defect in exiting stationary phase.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures






References
-
- Benoff B, Yang H, Lawson CL, Parkinson G, Liu J, Blatter E, Ebright YW, Berman HM, Ebright RH. Structural basis of transcription activation: the CAP-alpha CTD-DNA complex. Science. 2002;5586:1562–1566. - PubMed
-
- Borukhov S, Lee J, Laptenko O. Bacterial transcription elongation factors: new insights into molecular mechanism of action. Mol. Microbiol. 2005;5:1315–1324. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
