

Copsin, a novel peptide-based fungal antibiotic interfering with the peptidoglycan synthesis
- PMID: 25342741
- PMCID: PMC4263892
- DOI: 10.1074/jbc.M114.599878
Copsin, a novel peptide-based fungal antibiotic interfering with the peptidoglycan synthesis
Abstract
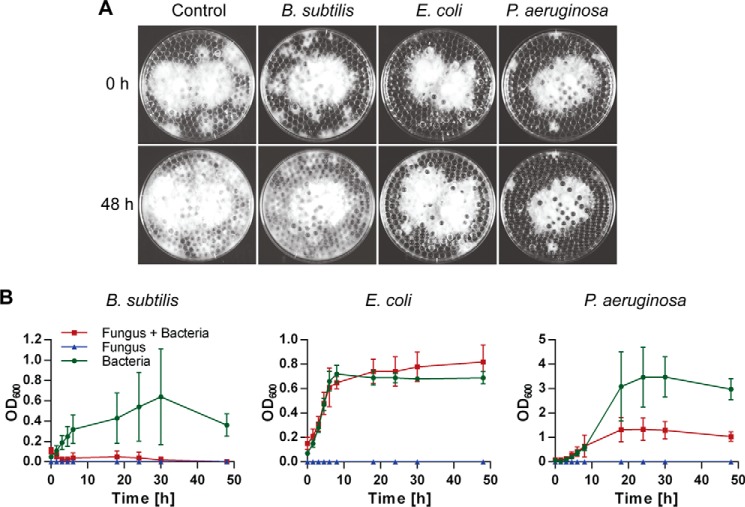
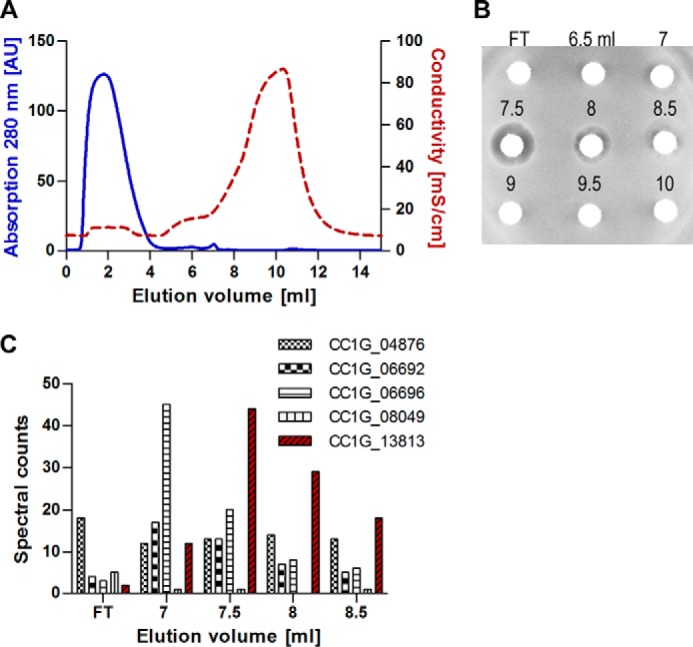
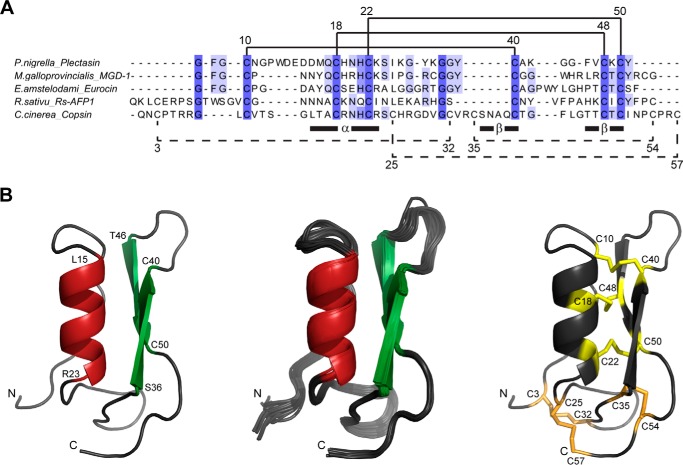
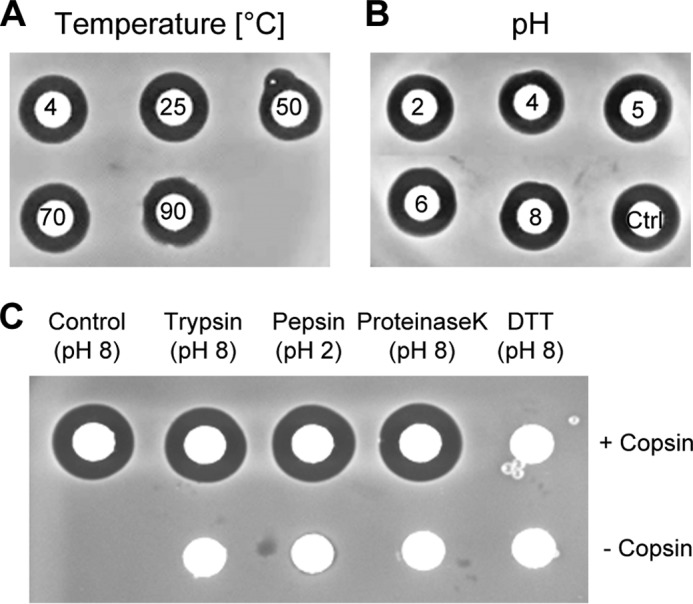
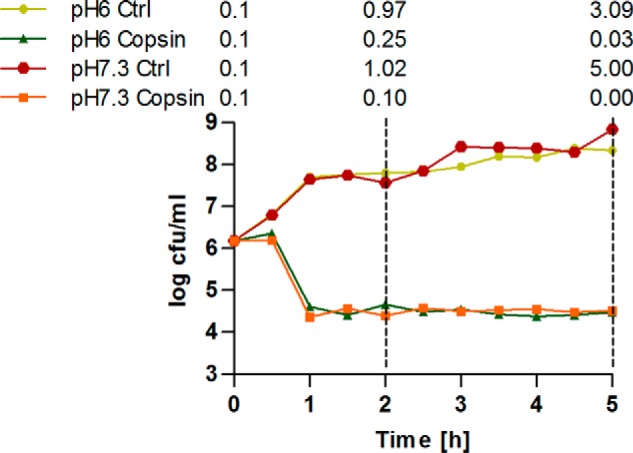
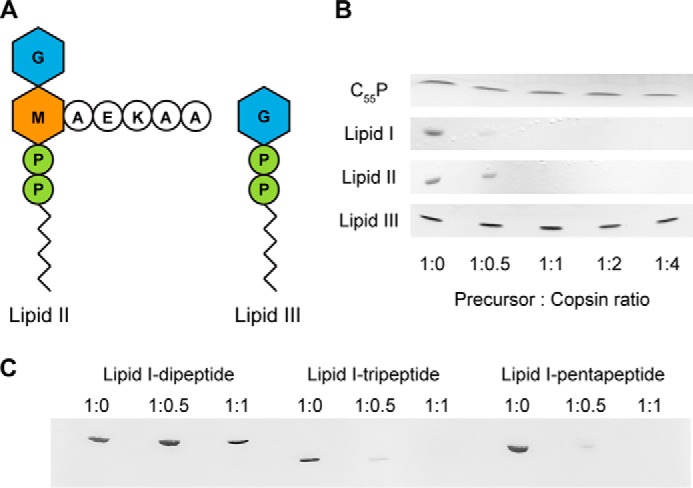
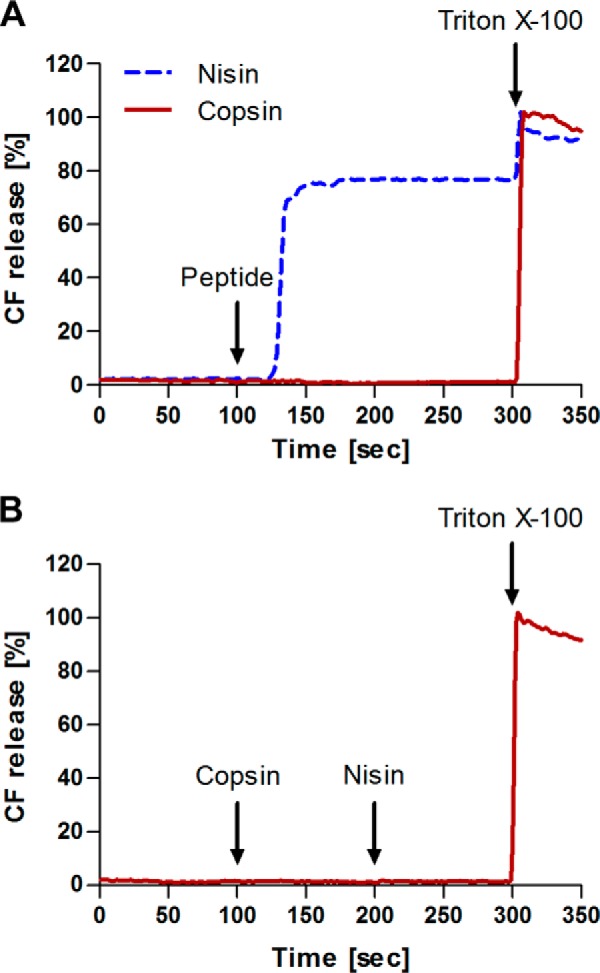
Fungi and bacteria compete with an arsenal of secreted molecules for their ecological niche. This repertoire represents a rich and inexhaustible source for antibiotics and fungicides. Antimicrobial peptides are an emerging class of fungal defense molecules that are promising candidates for pharmaceutical applications. Based on a co-cultivation system, we studied the interaction of the coprophilous basidiomycete Coprinopsis cinerea with different bacterial species and identified a novel defensin, copsin. The polypeptide was recombinantly produced in Pichia pastoris, and the three-dimensional structure was solved by NMR. The cysteine stabilized α/β-fold with a unique disulfide connectivity, and an N-terminal pyroglutamate rendered copsin extremely stable against high temperatures and protease digestion. Copsin was bactericidal against a diversity of Gram-positive bacteria, including human pathogens such as Enterococcus faecium and Listeria monocytogenes. Characterization of the antibacterial activity revealed that copsin bound specifically to the peptidoglycan precursor lipid II and therefore interfered with the cell wall biosynthesis. In particular, and unlike lantibiotics and other defensins, the third position of the lipid II pentapeptide is essential for effective copsin binding. The unique structural properties of copsin make it a possible scaffold for new antibiotics.
Keywords: Antibiotic Resistance; Antibiotics; Antimicrobial Peptide (AMP); Bacterial-Fungal Interaction (BFI); Fungal Secretome; Fungi; Lipid II.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Scherlach K., Graupner K., Hertweck C. (2013) Molecular bacteria-fungi interactions: effects on environment, food, and medicine. Annu. Rev. Microbiol. 67, 375–397 - PubMed
-
- Fleming A. (1929) On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. Br. J. Exp. Pathol. 10, 226–236 - PubMed
-
- Zasloff M. (2002) Antimicrobial peptides of multicellular organisms. Nature 415, 389–395 - PubMed
-
- Mygind P. H., Fischer R. L., Schnorr K. M., Hansen M. T., Sönksen C. P., Ludvigsen S., Raventós D., Buskov S., Christensen B., De Maria L., Taboureau O., Yaver D., Elvig-Jørgensen S. G., Sørensen M. V., Christensen B. E., Kjaerulff S., Frimodt-Moller N., Lehrer R. I., Zasloff M., Kristensen H. H. (2005) Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 437, 975–980 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
