

The Startle Disease Mutation E103K Impairs Activation of Human Homomeric α1 Glycine Receptors by Disrupting an Intersubunit Salt Bridge across the Agonist Binding Site
- PMID: 28174298
- PMCID: PMC5377815
- DOI: 10.1074/jbc.M116.767616
The Startle Disease Mutation E103K Impairs Activation of Human Homomeric α1 Glycine Receptors by Disrupting an Intersubunit Salt Bridge across the Agonist Binding Site
Abstract
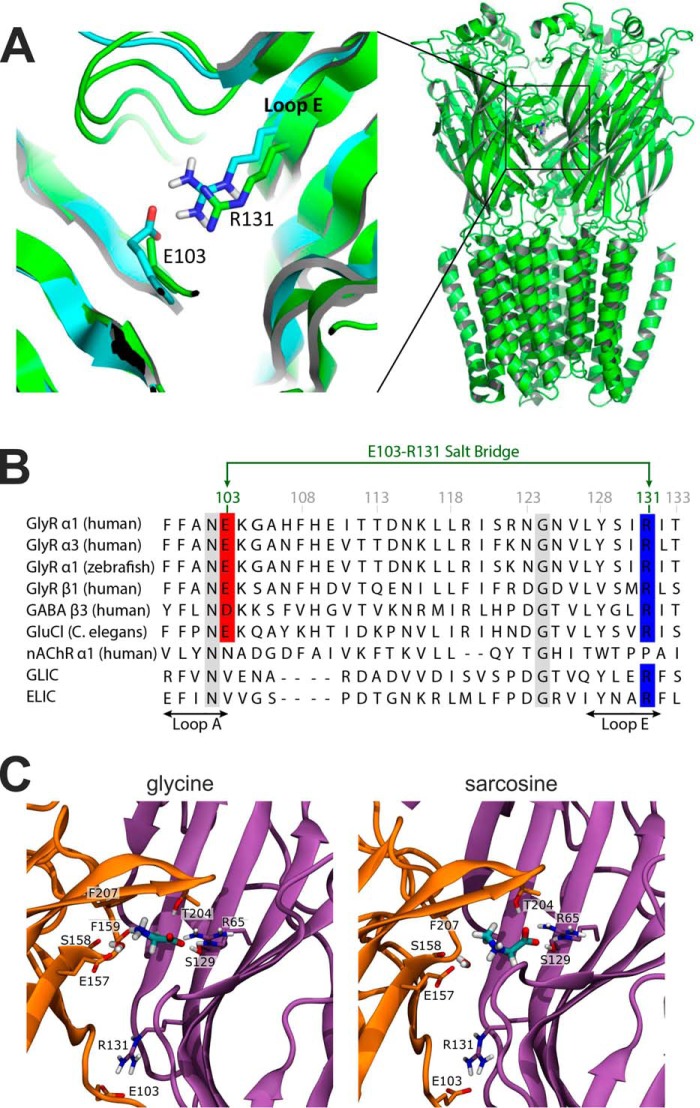
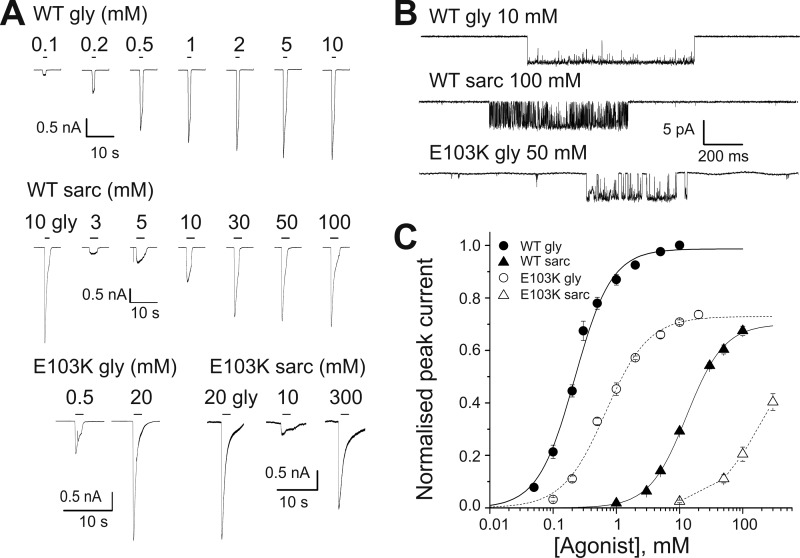
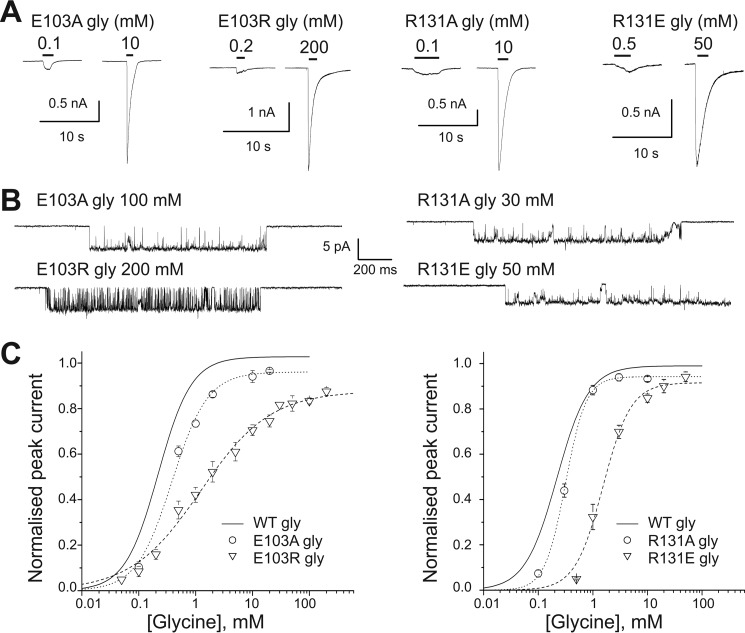
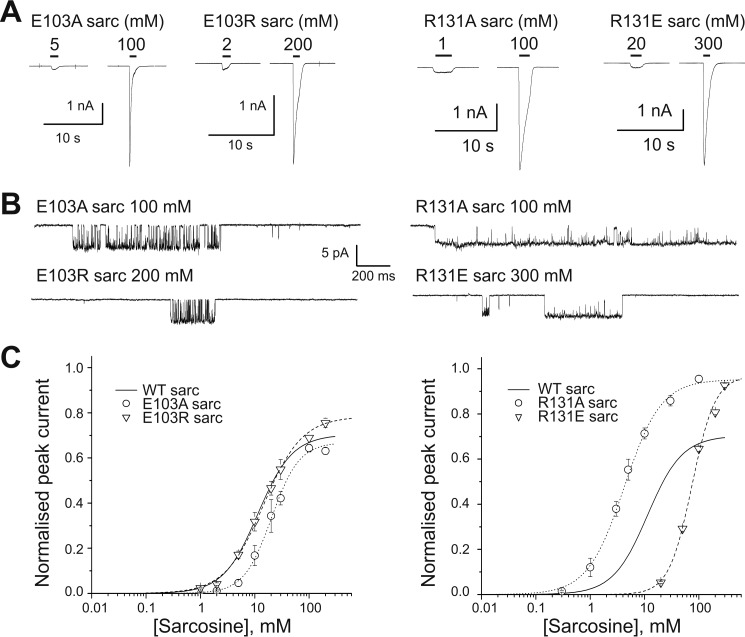
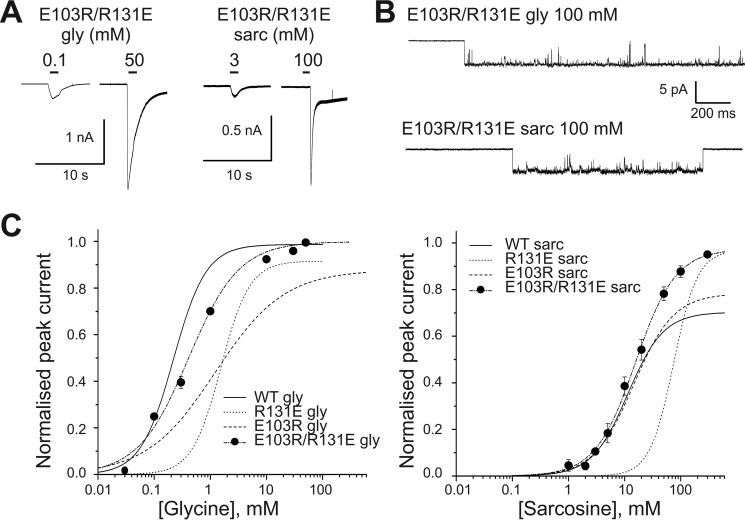
Glycine receptors (GlyR) belong to the pentameric ligand-gated ion channel (pLGIC) superfamily and mediate fast inhibitory transmission in the vertebrate CNS. Disruption of glycinergic transmission by inherited mutations produces startle disease in man. Many startle mutations are in GlyRs and provide useful clues to the function of the channel domains. E103K is one of few startle mutations found in the extracellular agonist binding site of the channel, in loop A of the principal side of the subunit interface. Homology modeling shows that the side chain of Glu-103 is close to that of Arg-131, in loop E of the complementary side of the binding site, and may form a salt bridge at the back of the binding site, constraining its size. We investigated this hypothesis in recombinant human α1 GlyR by site-directed mutagenesis and functional measurements of agonist efficacy and potency by whole cell patch clamp and single channel recording. Despite its position near the binding site, E103K causes hyperekplexia by impairing the efficacy of glycine, its ability to gate the channel once bound, which is very high in wild type GlyR. Mutating Glu-103 and Arg-131 caused various degrees of loss-of-function in the action of glycine, whereas mutations in Arg-131 enhanced the efficacy of the slightly bigger partial agonist sarcosine (N-methylglycine). The effects of the single charge-swapping mutations of these two residues were largely rescued in the double mutant, supporting the possibility that they interact via a salt bridge that normally constrains the efficacy of larger agonist molecules.
Keywords: agonists; efficacy; glycine receptor; homology modeling; ion channel; mutagenesis; patch clamp; potency; site-directed mutagenesis.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Galzi J.-L., and Changeux J.-P. (1995) Neuronal nicotinic receptors: molecular organization and regulations. Neuropharmacology 34, 563–582 - PubMed
-
- Beene D. L., Brandt G. S., Zhong W., Zacharias N. M., Lester H. A., and Dougherty D. A. (2002) Cation-π interactions in ligand recognition by serotonergic (5-HT3A) and nicotinic acetylcholine receptors: the anomalous binding properties of nicotine. Biochemistry 41, 10262–10269 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
