

How mutations in tRNA distant from the anticodon affect the fidelity of decoding
- PMID: 21378964
- PMCID: PMC3072312
- DOI: 10.1038/nsmb.2003
How mutations in tRNA distant from the anticodon affect the fidelity of decoding
Abstract
The ribosome converts genetic information into protein by selecting aminoacyl tRNAs whose anticodons base-pair to an mRNA codon. Mutations in the tRNA body can perturb this process and affect fidelity. The Hirsh suppressor is a well-studied tRNA(Trp) harboring a G24A mutation that allows readthrough of UGA stop codons. Here we present crystal structures of the 70S ribosome complexed with EF-Tu and aminoacyl tRNA (native tRNA(Trp), G24A tRNA(Trp) or the miscoding A9C tRNA(Trp)) bound to cognate UGG or near-cognate UGA codons, determined at 3.2-Å resolution. The A9C and G24A mutations lead to miscoding by facilitating the distortion of tRNA required for decoding. A9C accomplishes this by increasing tRNA flexibility, whereas G24A allows the formation of an additional hydrogen bond that stabilizes the distortion. Our results also suggest that each native tRNA will adopt a unique conformation when delivered to the ribosome that allows accurate decoding.
Figures
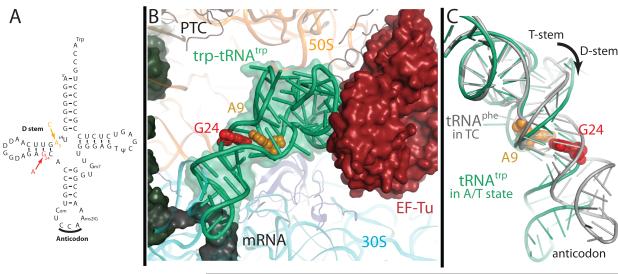
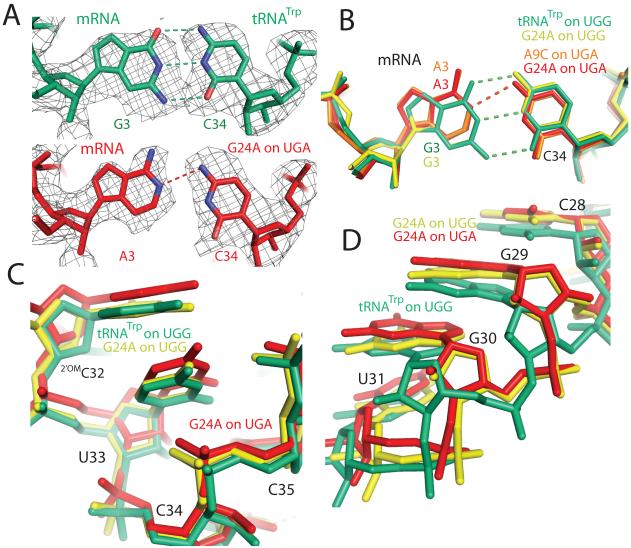
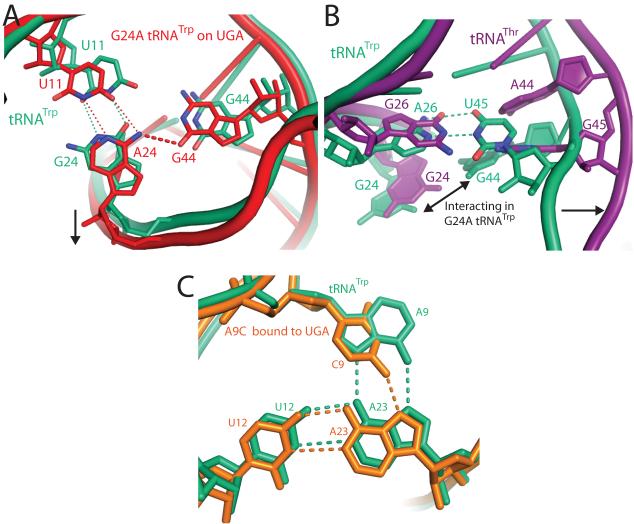
Comment in
-
Gene expression: Misreading the code.Nat Rev Mol Cell Biol. 2011 Apr;12(4):206. doi: 10.1038/nrm3095. Nat Rev Mol Cell Biol. 2011. PMID: 21427761 No abstract available.
References
-
- Hirsh D. Tryptophan tRNA of Escherichia coli. Nature. 1970;228:57. - PubMed
-
- Smith D, Yarus M. Transfer RNA structure and coding specificity. II. A D-arm tertiary interaction that restricts coding range. J Mol Biol. 1989;206:503–511. - PubMed
-
- Smith DW, Yarus M. Transfer RNA structure and coding specificity. I. Evidence that a D-arm mutation reduces tRNA dissociation from the ribosome. J Mol Biol. 1989;206:489–501. - PubMed
-
- Schultz DW, Yarus M. tRNA structure and ribosomal function. I. tRNA nucleotide 27-43 mutations enhance first position wobble. J Mol Biol. 1994;235:1381–1394. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
