

Structural and functional insights into DR2231 protein, the MazG-like nucleoside triphosphate pyrophosphohydrolase from Deinococcus radiodurans
- PMID: 21733847
- PMCID: PMC3162430
- DOI: 10.1074/jbc.M111.247999
Structural and functional insights into DR2231 protein, the MazG-like nucleoside triphosphate pyrophosphohydrolase from Deinococcus radiodurans
Abstract
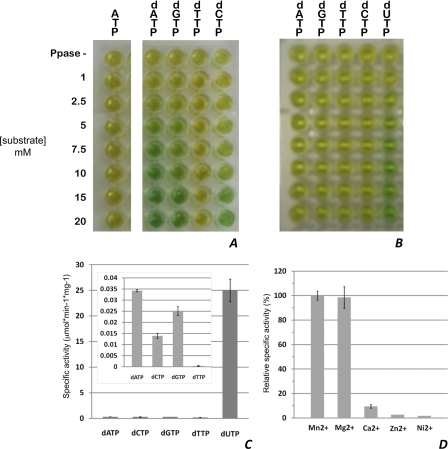
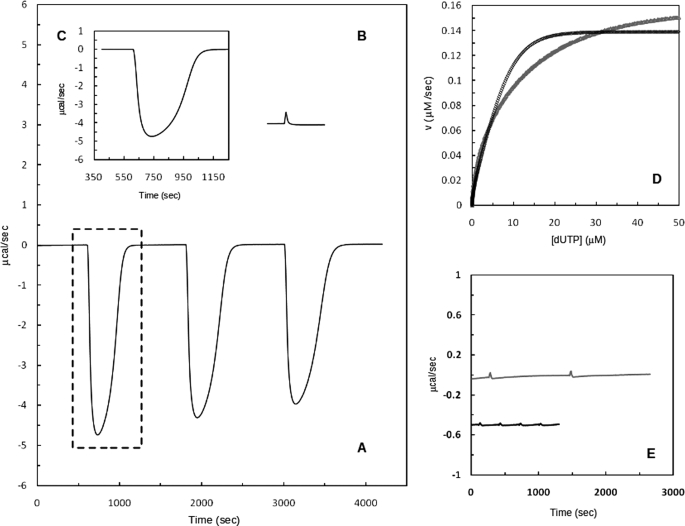
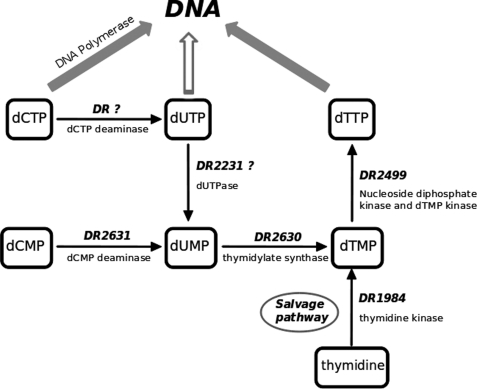
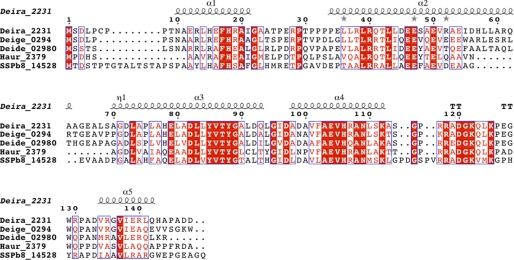
Deinococcus radiodurans is among the very few bacterial species extremely resistant to ionizing radiation, UV light, oxidizing agents, and cycles of prolonged desiccation. The proteome of D. radiodurans reflects the evolutionary pressure exerted by chronic exposure to (nonradioactive) forms of DNA and protein damage. A clear example of this adaptation is the overrepresentation of protein families involved in the removal of non-canonical nucleoside triphosphates (NTPs) whose incorporation into nascent DNA would promote mutagenesis and DNA damage. The three-dimensional structure of the DR2231 protein has been solved at 1.80 Å resolution. This protein had been classified as an all-α-helical MazG-like protein. The present study confirms that it holds the basic structural module characteristic of the MazG superfamily; two helices form a rigid domain, and two helices form a mobile domain and connecting loops. Contrary to what is known of MazG proteins, DR2231 protein shows a functional affinity with dUTPases. Enzymatic and isothermal calorimetry assays have demonstrated high specificity toward dUTP but an inability to hydrolyze dTTP, a typical feature of dUTPases. Co-crystallization with the product of hydrolysis, dUMP, in the presence of magnesium or manganese cations, suggests similarities with the dUTP/dUDP hydrolysis mechanism reported for dimeric dUTPases. The genome of D. radiodurans encodes for all enzymes required for dTTP synthesis from dCMP, thus bypassing the need of a dUTPase. We postulate that DR2231 protein is not essential to D. radiodurans and rather performs "house-cleaning" functions within the framework of oxidative stress response. We further propose DR2231 protein as an evolutionary precursor of dimeric dUTPases.
Figures
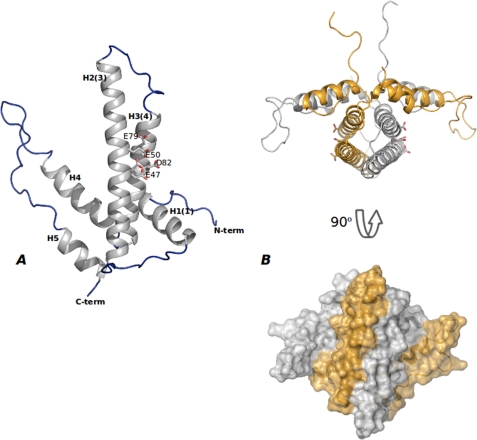

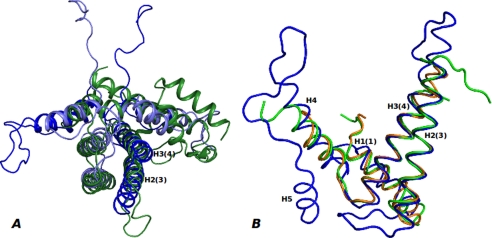
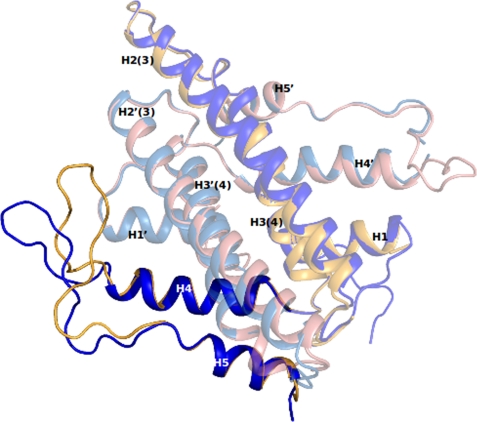
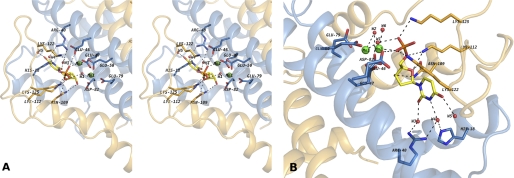
References
-
- Bessman M. J., Frick D. N., O'Handley S. F. (1996) J. Biol. Chem. 271, 25059–25062 - PubMed
-
- Koonin E. V., Aravind L., Galperin M. Y. (2000) in Bacterial Stress Responses (Storz G., Hengge-Aronis R. eds) pp. 417–444, American Society for Microbiology Press, Washington, D. C
-
- Makarova K. S., Aravind L., Daly M. J., Koonin E. V. (2000) Genetica 108, 25–34 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
