

The Structure of the Transcriptional Repressor KstR in Complex with CoA Thioester Cholesterol Metabolites Sheds Light on the Regulation of Cholesterol Catabolism in Mycobacterium tuberculosis
- PMID: 26858250
- PMCID: PMC4817160
- DOI: 10.1074/jbc.M115.707760
The Structure of the Transcriptional Repressor KstR in Complex with CoA Thioester Cholesterol Metabolites Sheds Light on the Regulation of Cholesterol Catabolism in Mycobacterium tuberculosis
Abstract
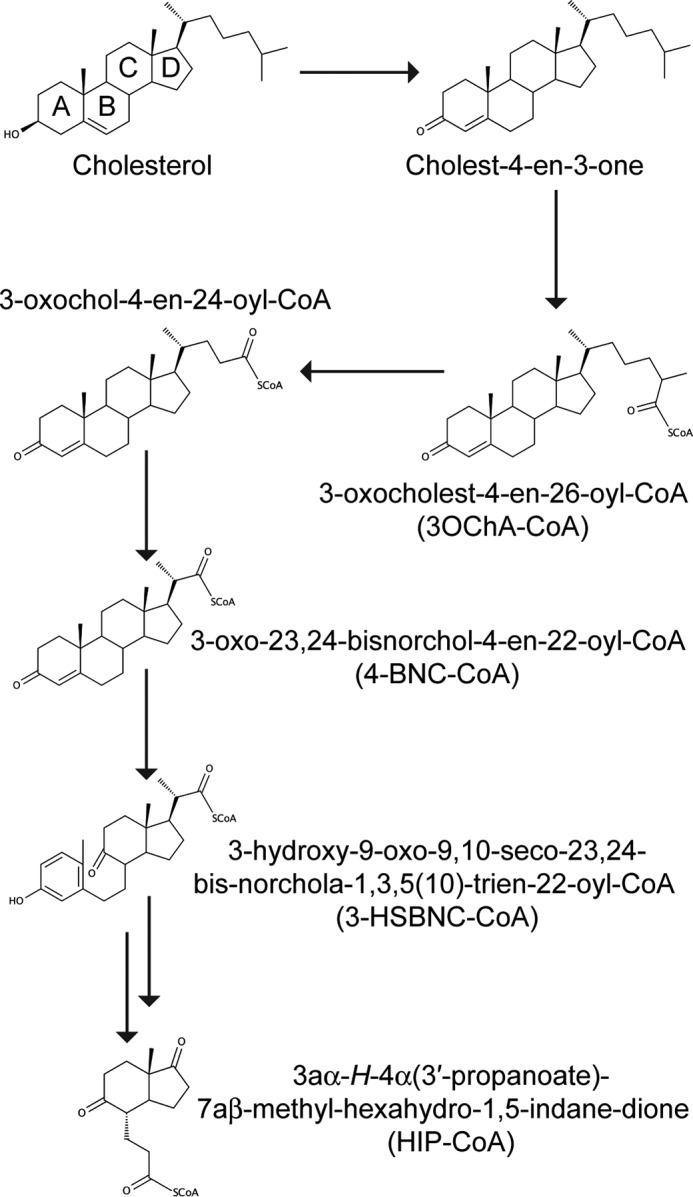
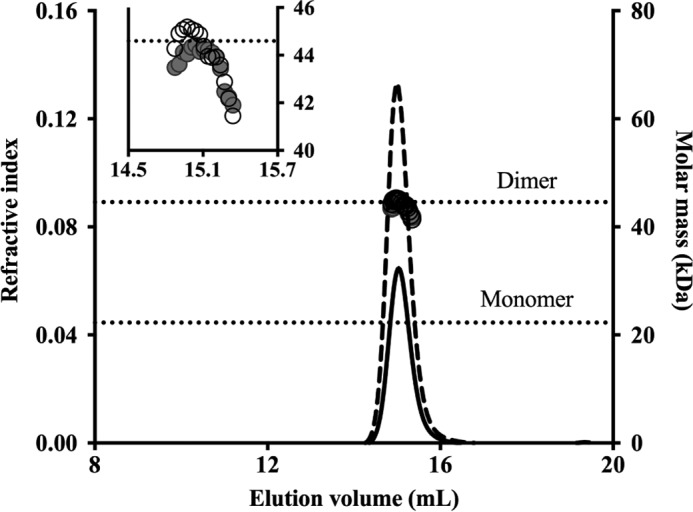
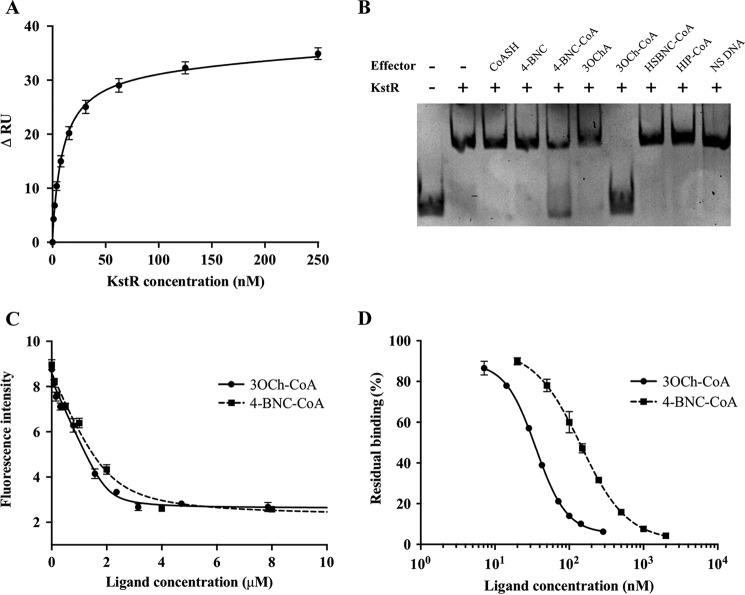
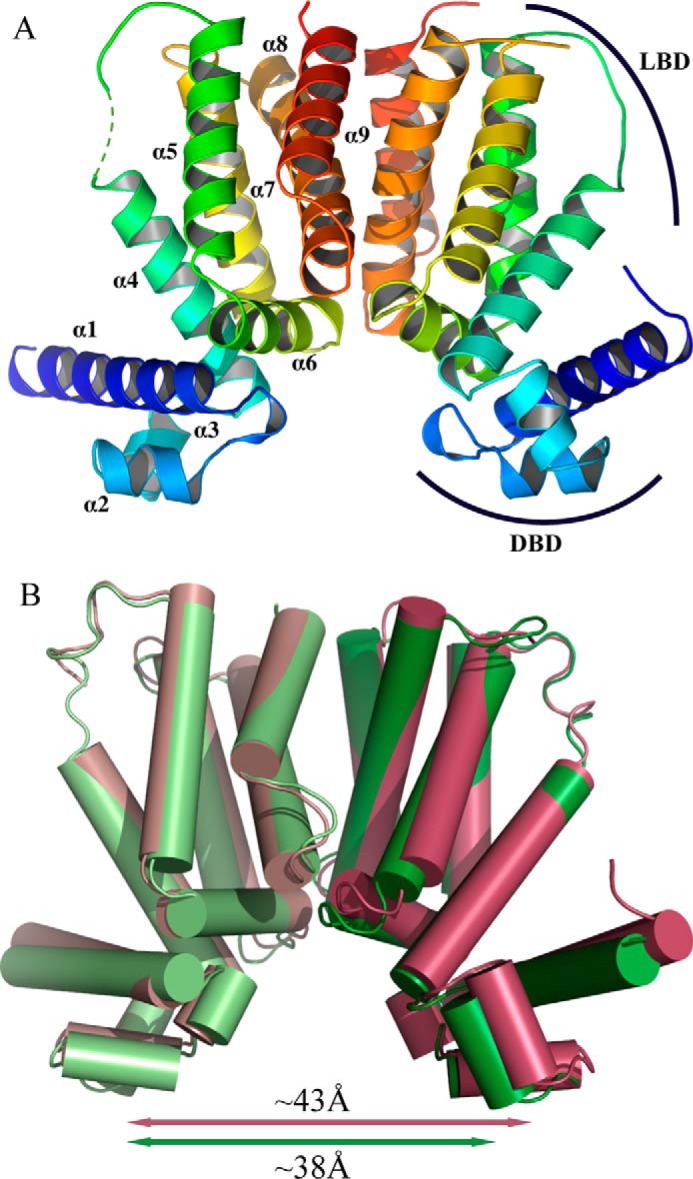
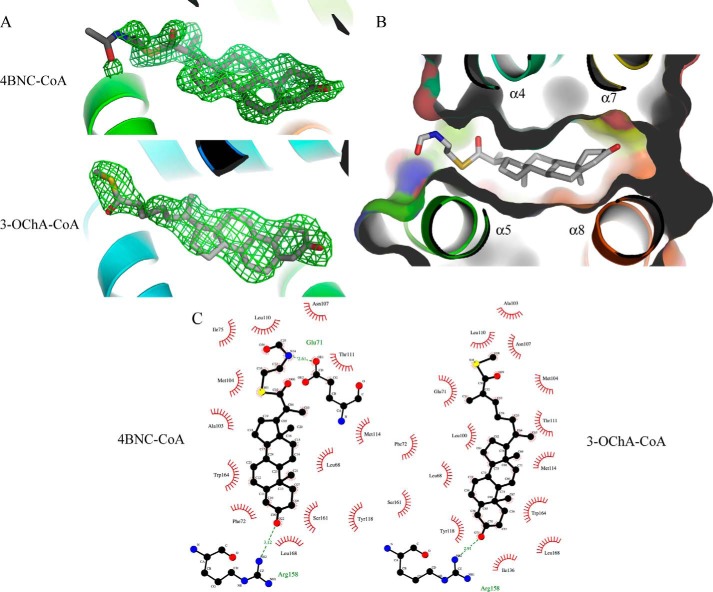
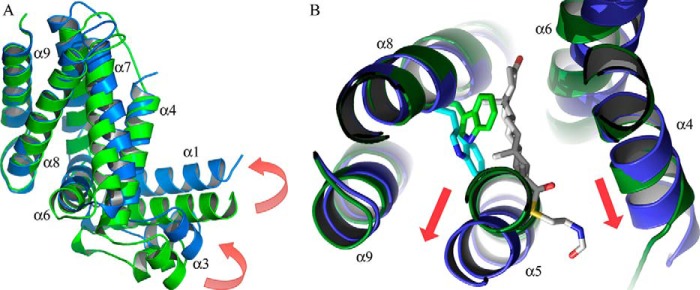
Cholesterol can be a major carbon source forMycobacterium tuberculosisduring infection, both at an early stage in the macrophage phagosome and later within the necrotic granuloma. KstR is a highly conserved TetR family transcriptional repressor that regulates a large set of genes responsible for cholesterol catabolism. Many genes in this regulon, includingkstR, are either induced during infection or are essential for survival ofM. tuberculosis in vivo In this study, we identified two ligands for KstR, both of which are CoA thioester cholesterol metabolites with four intact steroid rings. A metabolite in which one of the rings was cleaved was not a ligand. We confirmed the ligand-protein interactions using intrinsic tryptophan fluorescence and showed that ligand binding strongly inhibited KstR-DNA binding using surface plasmon resonance (IC50for ligand = 25 nm). Crystal structures of the ligand-free form of KstR show variability in the position of the DNA-binding domain. In contrast, structures of KstR·ligand complexes are highly similar to each other and demonstrate a position of the DNA-binding domain that is unfavorable for DNA binding. Comparison of ligand-bound and ligand-free structures identifies residues involved in ligand specificity and reveals a distinctive mechanism by which the ligand-induced conformational change mediates DNA release.
Keywords: CoA thioester; Mycobacterium tuberculosis; bacterial metabolism; cholesterol metabolism; protein structure; transcription repressor.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- World Health Organization (2015) Global Tuberculosis Report 2015, World Health Organization, Geneva
-
- Cole S. T., Brosch R., Parkhill J., Garnier T., Churcher C., Harris D., Gordon S. V., Eiglmeier K., Gas S., Barry C. E. 3rd, Tekaia F., Badcock K., Basham D., Brown D., Chillingworth T., et al. (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393, 537–544 - PubMed
-
- Yam K. C., D'Angelo I., Kalscheuer R., Zhu H., Wang J.-X., Snieckus V., Ly L. H., Converse P. J., Jacobs W. R. Jr., Strynadka N., and Eltis L. D. (2009) Studies of a ring-cleaving dioxygenase illuminate the role of cholesterol metabolism in the pathogenesis of Mycobacterium tuberculosis. PLoS Pathog. 5, e1000344. - PMC - PubMed
-
- Brzostek A., Dziadek B., Rumijowska-Galewicz A., Pawelczyk J., and Dziadek J. (2007) Cholesterol oxidase is required for virulence of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 275, 106–112 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
