

The bacterial deubiquitinase Ceg23 regulates the association of Lys-63-linked polyubiquitin molecules on the Legionella phagosome
- PMID: 31907282
- PMCID: PMC7008378
- DOI: 10.1074/jbc.RA119.011758
The bacterial deubiquitinase Ceg23 regulates the association of Lys-63-linked polyubiquitin molecules on the Legionella phagosome
Abstract
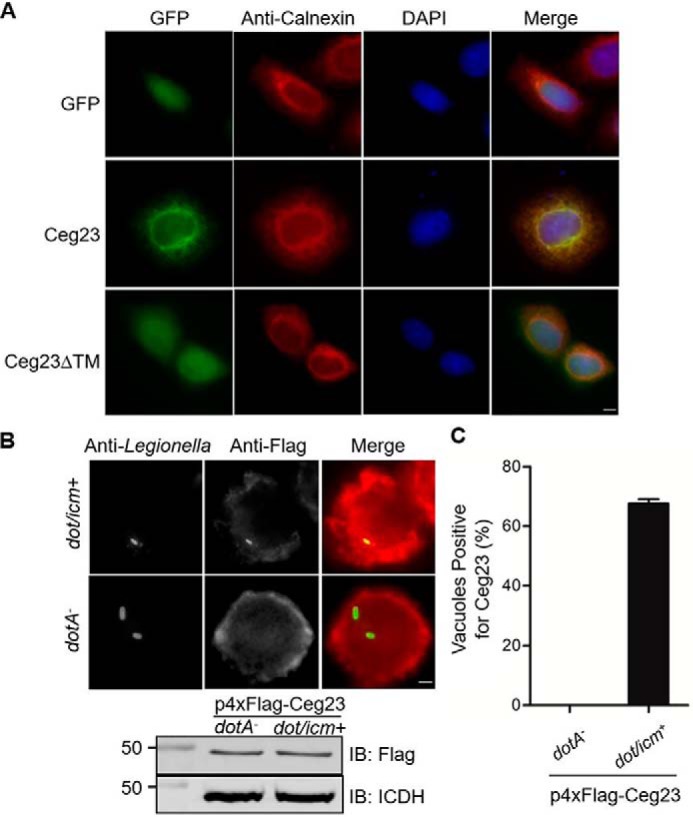
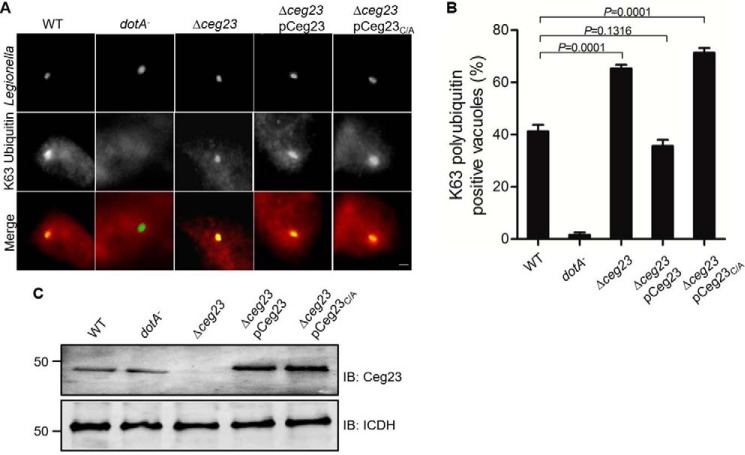
Legionella pneumophila is the causative agent of the lung malady Legionnaires' disease, it modulates host function to create a niche termed the Legionella-containing vacuole (LCV) that permits intracellular L. pneumophila replication. One important aspect of such modulation is the co-option of the host ubiquitin network with a panel of effector proteins. Here, using recombinantly expressed and purified proteins, analytic ultracentrifugation, structural analysis, and computational modeling, along with deubiquitinase (DUB), and bacterial infection assays, we found that the bacterial defective in organelle trafficking/intracellular multiplication effector Ceg23 is a member of the ovarian tumor (OTU) DUB family. We found that Ceg23 displays high specificity toward Lys-63-linked polyubiquitin chains and is localized on the LCV, where it removes ubiquitin moieties from proteins ubiquitinated by the Lys-63-chain type. Analysis of the crystal structure of a Ceg23 variant lacking two putative transmembrane domains at 2.80 Å resolution revealed that despite very limited homology to established members of the OTU family at the primary sequence level, Ceg23 harbors a catalytic motif resembling those associated with typical OTU-type DUBs. ceg23 deletion increased the association of Lys-63-linked polyubiquitin with the bacterial phagosome, indicating that Ceg23 regulates Lys-63-linked ubiquitin signaling on the LCV. In summary, our findings indicate that Ceg23 contributes to the regulation of the association of Lys-63 type polyubiquitin with the Legionella phagosome. Future identification of host substrates targeted by Ceg23 could clarify the roles of these polyubiquitin chains in the intracellular life cycle of L. pneumophila and Ceg23's role in bacterial virulence.
Keywords: Legionella pneumophila; bacteria; bacterial pathogenesis; deubiquitylation (deubiquitination); effector protein; ovarian-tumor family deubiquitinases; polyubiquitin chain; structural biology; type IV secretion system.
© 2020 Ma et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
