

Structural basis for the coupling between activation and inactivation gates in K(+) channels
- PMID: 20613845
- PMCID: PMC3033755
- DOI: 10.1038/nature09136
Structural basis for the coupling between activation and inactivation gates in K(+) channels
Abstract
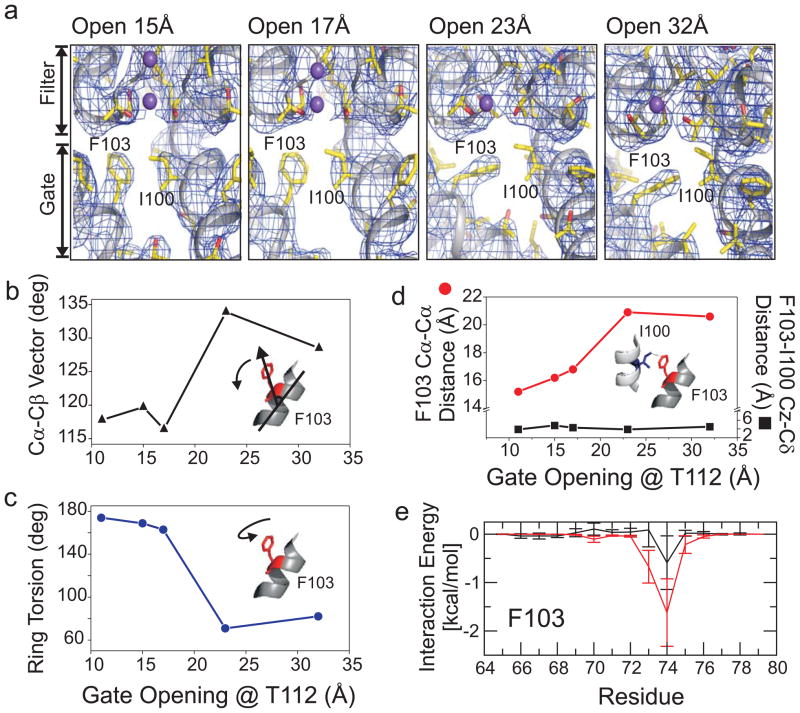
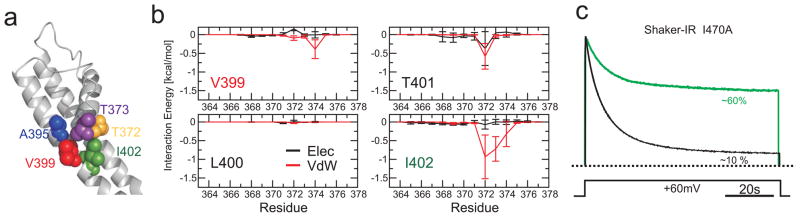
The coupled interplay between activation and inactivation gating is a functional hallmark of K(+) channels. This coupling has been experimentally demonstrated through ion interaction effects and cysteine accessibility, and is associated with a well defined boundary of energetically coupled residues. The structure of the K(+) channel KcsA in its fully open conformation, in addition to four other partial channel openings, richly illustrates the structural basis of activation-inactivation gating. Here, we identify the mechanistic principles by which movements on the inner bundle gate trigger conformational changes at the selectivity filter, leading to the non-conductive C-type inactivated state. Analysis of a series of KcsA open structures suggests that, as a consequence of the hinge-bending and rotation of the TM2 helix, the aromatic ring of Phe 103 tilts towards residues Thr 74 and Thr 75 in the pore-helix and towards Ile 100 in the neighbouring subunit. This allows the network of hydrogen bonds among residues Trp 67, Glu 71 and Asp 80 to destabilize the selectivity filter, allowing entry to its non-conductive conformation. Mutations at position 103 have a size-dependent effect on gating kinetics: small side-chain substitutions F103A and F103C severely impair inactivation kinetics, whereas larger side chains such as F103W have more subtle effects. This suggests that the allosteric coupling between the inner helical bundle and the selectivity filter might rely on straightforward mechanical deformation propagated through a network of steric contacts. Average interactions calculated from molecular dynamics simulations show favourable open-state interaction-energies between Phe 103 and the surrounding residues. We probed similar interactions in the Shaker K(+) channel where inactivation was impaired in the mutant I470A. We propose that side-chain rearrangements at position 103 mechanically couple activation and inactivation in KcsA and a variety of other K(+) channels.
Figures


References
-
- Baukrowitz T, Yellen G. Use-dependent blockers and exit rate of the last ion from the multi-ion pore of a K+ channel. Science. 1996;271:653–656. - PubMed
-
- Lopez-Barneo J, Hoshi T, Heinemann SH, Aldrich RW. Effects of external cations and mutations in the pore region on C-type inactivation of Shaker potassium channels. Receptors Channels. 1993;1:61–71. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
